Микроорганизмы, объекты биотехнологии
Основа микробиологии: Микроорганизмы

Вирусы
ВВЕДЕНИЕ. Вирусы не обладают собственным обменом веществ. Репликация генетического материала вирусов – ДНК или РНК – происходит с помощью клеток-хозяев. Вне клетки-хозяина вирус представляет собой нуклеиновую кислоту, одетую белковой оболочкой (капсидом). Такое состояние вируса называется нуклеокапсидом или вирионом. Вирусы могут инфицировать большинство живых организмов, однако они проявляют высокую специфичность по отношению к клеткам-хозяевам, выбирая определенные ткани или клетки в организме. Существует несколько способов классификации вирусов: по типам клеток-хозяев, по морфологическим признакам, по генетическому материалу (ДНК или РНК) или по строению капсида. Вирусы широко используются в биотехнологии для получения многокомпонентных вакцин, а также для разработки различных векторов, например, для генной терапии или для экспрессии генов в культурах клеток.
ВИРУСЫ В ЭКСПЕРИМЕНТАХ С ЖИВОТНЫМИ. Первые эксперименты по клонированию животных клеток были проведены в 1979 г. с использованием вектора на основе вируса обезьян (SV40). Вирус проникает в клетку, а затем его развитие протекает по литическому или лизогенному пути. Геном вируса (5,2 т.п.н.) содержит так называемые «ранние гены», кодирующие белки, необходимые для репликации ДНК, и «поздние гены», продукты которых участвуют в синтезе капсида. Векторы на основе SV40 содержат вирусные регуляторные элементы: точку начала репликации, промоторный участок, а также терминатор транскрипции (сайт полиаденилирования). Для трансфекции клеток мышей используют конструкции на основе папилломавируса крупного рогатого скота (BPV). При инфекции они ведут себя как высококопийные плазмиды, и ДНК-копии передаются при клеточном делении дочерним клеткам. Ослабленные ретровирусы, аденовирусы, а также вирус герпеса применяются для генной терапии. Геном ретровирусов (например, ВИЧ) представляет собой РНК. Ретровирусы инфицируют делящиеся клетки, при этом обратная транскриптаза, кодированная в вирусном геноме, обеспечивает синтез кДНК-копии вирусного РНК-генома. Эта кДНК встраивается в геном хозяина и использует его сильные промоторы для синтеза белков капсида и вирусной мРНК. В экспериментальной генной терапии успешно используются ретровирусы с дефектами репликации, однако размер ДНК, доставляемой с помощью вируса, невелик. В отличие от ретровирусов аденовирусы могут быть использованы для упаковки крупных молекул ДНК (до 28 т.п.н.). Аденовирусы инфицируют как делящиеся, так и неделящиеся клетки, однако их ДНК не встраивается в геном хозяина. Известен случай, когда использование аденовирусов вызвало неожиданную иммунную реакцию и привело к смерти 18-летнего пациента. С тех пор использование аденовирусов в медицине практически прекращено. В настоящее время изучается возможность использования векторов на основе вируса простого герпеса Нerpes simplex для генной терапии таких нервных расстройств, как болезни Паркинсона или Альцгеймера. Вирус Нerpes simplex обладает большим ДНК-геномом (152 т.п.н.), следовательно, в него можно встраивать крупные фрагменты ДНК.
ВИРУСЫ В ЭКСПЕРИМЕНТАХ С РАСТЕНИЯМИ. Большинство вирусов растений имеют РНК-геном.Известны лишь две группы ДНК-содержащих вирусов, которые могут инфицировать высшие растения.У каулимовирусов (или колимовирусов) спектр клеток-хозяев очень узкий:они поражают только представителей семейства крестоцветных – свеклу и некоторые сорта капусты. Каулимовирусы имеют очень маленький капсид, поэтому их собственный геном и чужеродная упакованнаяДНК очень невелики. Геминивирусы инфицируют такие важные сельскохозяйственные культуры, как кукуруза и пшеница, поэтому их использование сопряжено с высокой степенью риска. Кроме того, приинфекции в геноме геминивирусов происходят множественные перестройки, в том числе и делецииДНК, поэтому часто возникают проблемы с экспрессией встроенных фрагментов ДНК.
БАКУЛОВИРУСЫ. Эти вирусы заражают насекомых, но безопасны для позвоночных. В результате вирусной инфекции в клетке начинается синтез кристаллического белка полигедрина (полиэдрина), а синтез более половины белков клетки-хозяина подавляется. Промотор полигедрина используется при создании векторов для гетерологичной экспрессии в клеточной культуре Spodoptera (род бабочек). Премущество заключается в том, что посттрансляционное гликозилирование в такой системе аналогично таковому у позвоночных. В настоящее время перечисленные системы экспрессии используются только в лабораторных исследованиях.


Бактериофаги
ВВЕДЕНИЕ. Бактериофагами (фагами) называются вирусы, которые инфицируют бактерии. Устойчивость к фаговой инфекции – один из важных критериев при получении штаммов-суперпродуцентов. В генетической инженерии бактериофаги используются для получения век торов и промоторов для клонирования, для секвенирования и для создания геномных или белковых библиотек. Чаще всего в качестве клеток-хозяев для клонирования используют клетки E. coli, поэтому бактериофаги (фаги λ, М13, Qβ, Т-фаг), заражающие эти бактерии, вызывают наибольший интерес.
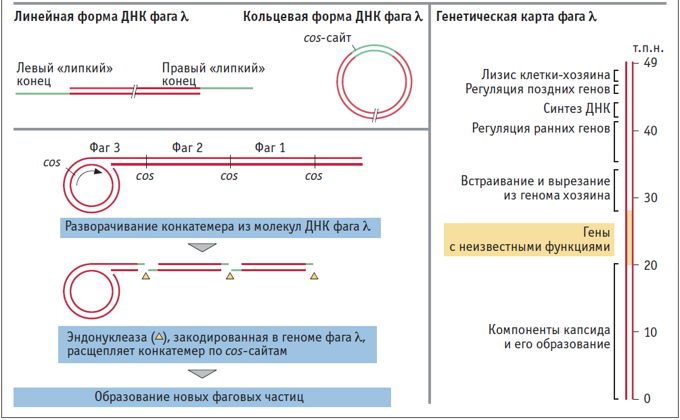
ФАГ λ инфицирует клетки E. coli. Как и другие представители умеренных фагов, после заражения фаг λ может развиваться по одному из двух путей: литическому или лизогенному. При литическом росте геном фага, который представлен линейной двунитевой ДНК размером 48,5 т.п.н., многократно реплицируется вне хромосомы хозяина, а затем фаговые частицы высвобождаются из клетки, лизируя ее. Если реализуется лизогенный путь развития фага, его ДНК (размером
1% хромосомной ДНК E. coli) встраивается в геном хозяина и реплицируется вместе с ним. Бактерии, содержащие интегрированный геном фага (профага), называются лизогенными. Повышение температуры, УФ-облучение или другой стресс приводит к высвобождению профага из генома E. coli и лизису клетки. В ДНК фага λ имеются так называемые cos-сайты – одноцепочечные 5′-концевые участки длиной 12 нуклеотидов, способные к комплементарному взаимодействию. После проникновения фага в клетку cos-сайты замыкаются, и образуется кольцевая молекула ДНК, репликация которой происходит по принципу «катящегося кольца». При этом образуются конкатемеры – последовательно соединенные копии фагового генома. Эндонуклеаза А – продукт гена А – расщепляет такую длинную молекулу по cos-сайтам, а затем отдельные молекулы фаговой ДНК пакуются в капсиды. На основе фага λ сконструировано множество векторов: например, космиды, используемые при создании геномных библиотек, или семейство λ-векторов, в которое входит вектор λEMBL4, индукция генов которого происходит при повышении температуры.
ФАГ М13 также инфицирует клетки E. coli, однако по строению этот вирус значительно отличается от фага λ. Геном фага М13 представляет собой одну молекулу одноцепочечной ДНК размером 6,4 т.п.н. После проникновения ДНК фага в клетку E. coli происходит синтез комплементарной цепи ДНК, и образовавшаяся двухцепочечная фаговая ДНК не встраивается в геном хозяина, а реплицируется в цитоплазме. Затем зрелые одноцепочечные молекулы ДНК фага М13 выходят из клетки, покрываясь капсидом (
1000 фаговых частиц на клетку). Инфицированные клетки при этом не погибают и при делении передают фаговую ДНК дочерним клеткам (
100 молекул на клетку). Особенности жизненного цикла бактериофага М13 используются при получении исследуемых генов в виде одноцепочечной ДНК, например для секвенирования ДНК, а также при сайт-направленном мутагенезе с применением ПЦР.
Т-ФАГИ разделены на семь типов. В генетической инженерии широко используются два фермента, кодированные в геноме Т-фагов: ДНК-лигаза фага Т4 – фермент, соединяющий «липкие» и «тупые» концы двух фрагментов ДНК, и ДНК-полимераза фага Т7, применяющаяся для секвенирования ДНК по методу Сэнгера–Коулсона. Промотор РНК-полимеразы фага Т7 часто встраивают в векторы для экспрессии белков в клетках E. coli.
ФАГИ ДРУГИХ БАКТЕРИЙ. Среди более чем тысячи охарактеризованных фагов более 300 инфицируют энтеробактерии, 230 заражают бактериококки, а 150 – актиномицеты и бациллы. Для представителей рода Pseudomonas описаны более 100, а для бактерий рода Lactobacillus – 40 фагов. По строению и физиологии эти вирусы отличаются от фагов, специфических для E. coli. При производстве молочных продуктов особенно важна защита от фагов, инфицирующих бактерии рода Lactobacillus, так как они могут присутствовать в стартовых культурах. Как правило, стартовые культуры представлены генетически модифицированными штаммами, устойчивыми к фаговой инфекции. Такая устойчивость обусловлена экспрессией белковых продуктов, кодированных в плазмиде. В результате действия этих белков могут нарушаться процессы внедрения вируса в клетку или его репликации. Среди пяти изученных групп вирусов бактерий рода Bacillus фаги Æ105 и SPO2 широко используется для трансформации, а фаг PBS1 – при построении карты генома Bacillus subtilis. Фаг М3112 часто служит для трансформации бактерий рода Pseudomonas, а фаги SV1 и ÆC31 – для введения ДНК в клетки Streptomyces.


Микроорганизмы
ВВЕДЕНИЕ. Многие ключевые реакции круговорота веществ в природе осуществляются только микроорганизмами. В частности, микроорганизмам принадлежит ведущая роль в осуществлении процессов распада. Эти процессы особенно важны для высших организмов, поэтому в природе часто имеют место симбиотические отношения между высшими организмами и микроорганизмами. В качестве примеров можно привести лишайники (симбиоз между грибами и водорослями), бактерии в рубце у крупного рогатого скота или кишечную флору млекопитающих. В то же время некоторые микроорганизмы являются возбудителями различных заболеваний. Непатогенные микроорганизмы широко используются в биотехнологии: при получения таких важных продуктов, как лимонная кислота, антибиотики, ксантановые смолы и применяемые в научных и производственных целях ферменты, а также при аэробной или анаэробной очистке сточных вод, воздуха и почвы и в синтезе рекомбинантных белков. В силу своего относительно простого строения микроорганизмы часто служат в качестве модельных организмов при изучении биохимических, генетических и физиологических процессов. Разработано много методик проведения мутагенеза, и преимущество микроорганизмов для таких экспериментов заключается в их сравнительно коротком жизненном цикле. Раньше классификация микроорганизмов была основана на клеточном строении: их делили на прокариотические и эукариотические; однако согласно современным представлениям, среди прокариот выделены также археи и эубактерии (около 6000 полностью охарактеризованных штаммов).
ЭУБАКТЕРИИ, или истинные бактерии, – это одноклеточные организмы размером около 1 мкм, размножающиеся делением. Как и все прокариоты, эубактерии не имеют клеточного ядра. ДНК эубактерии называется нуклеоидом. Эубактерии часто содержат нехромосомные ДНК, например плазмиды, в которых хранится часть генетического материала. Плазмиды могут распространяться в результате горизонтального обмена генами – чрезвычайно важного процесса, обеспечивающего естественную эволюцию метаболизма бактерий, в том числе образование штаммов, устойчивых к антибиотикам. В зависимости от строения клеточной стенки бактерии делятся на грамположительные и грамотрицательные, при этом грамотрицательные бактерии имеют более сложно устроенную клеточную стенку, чем бактерии, окрашивающиеся по методу Грама (грамположительные). Клетки многих бактерий покрыты слизистой оболочкой, а также имеют выросты, позволяющие им передвигаться. В цитоплазме бактериальных клеток могу накапливаться запасные вещества, например полигидроксимасляная кислота или полифосфаты. Благодаря разнообразию обмена веществ представители группы эубактерий встречаются в любой природной среде, этому также способствуют своеобразные пути эволюционирования белков и кофакторов эубактерий. Например, пурпурная мембрана галобактерий обладает некоторыми функциональными свойствами (в том числе способностью к фотосинтезу), позволяющими считать этих бактерий древнейшими предшественниками высших организмов.
АРХЕБАКТЕРИИ (АРХЕИ). Считается, что археи существовали сотни миллионов лет до нашей эры и что это одна из самых древних форм жизни на Земле. Для большинства архей характерен анаэробный обмен веществ (т. е. они могут существовать в отсутствие кисло рода), и они часто встречаются в так называемых экстремальных средах. Например, только представители архей осуществляют превращение уксусной кис лоты в метан – реакция, лежащая в основе биотехнологической очистки сточных вод. От эубактерий археи отличаются по структурным и генетическим признакам. Например, клеточная стенка архей построена не из фосфолипидов, а из эфиров глицерина. В связи с экстремальными условиями обитания архей их ферменты часто обладают специфическими свойствами, и эти особенности используются в биотехнологии. Так, ДНК-полимераза Pyrococcus furiosus, обитающей в глубоководных озерах, применяется для ПЦР в тех случаях, когда необходима повышенная точность интерполяции нуклеотидов.
ДРОЖЖИ И ГРИБЫ. Все дрожжи и грибы (около 70 000 штаммов) – эукариоты. В отличие от прокариот их клеточная стенка построена из хитина или, реже, из целлюлозы. Практически все грибы – гетеротрофы и осуществляют аэробное дыхание. В основе классификации грибов лежит способ их размножения. Вегетативное тело грибов (мицелий) состоит из системы ветвящихся нитей (гиф). Наиболее распространено бесполое размножение, которое осуществляется спорами или в результате почкования. Половой процесс у низших грибов (фикомицетов) происходит с участием гамет, а у высших грибов для этой цели формируются разнообразные плодовые тела, например сумки у аскомицетов или базидии у базидиомицетов. Форма плодового тела также признак, используемый при классификации грибов.


Бактерии
ВВЕДЕНИЕ. Бактерии – группа микроорганизмов, различающихся по множеству морфологических, биохимических и генетических признаков. В связи с этим возможны различные способы классификации бактерий. В настоящее время по коду международной номенклатуры бактерий (ICNB) зарегистрировано около 6 000 штаммов микроорганизмов. Молекулярно-генетический анализ рибосомной РНК из природных сред позволяет предположить, что количество еще не изученных бактерий значительно превышает количество зарегистрированных.
ЭУБАКТЕРИИ. Классическое определение бактерий основывалось на морфологических признаках: даже с помощью светового микроскопа можно различать палочки, кокки, спириллы, объединенные между собой клетки (колонии, филаменты), а также структурные особенности спор и гиф. Для дальнейшей классификации бактерий используется специфическое окрашивание клеток. Реакция клеток на окрашивание по Граму отражает особенности строения клеточной стенки: грамположительные бактерии имеют массивную многослойную клеточную стенку, построенную из муреина, под которой располагается плазматическая мембрана. Клеточное содержимое грамотрицательных бактерий одето внутренней и внешней клеточными мембранами, между которыми находится периплазматическое пространство. Поверх внешней мембраны располагается однослойная клеточная стенка, построенная из муреина и сложных липополисахаридов. Более детальная классификация бактерий возможна при изучении их физиологических и биохимических особенностей. Можно выделить несколько основных критериев для классификации.
Отношение к кислороду: рост бактерий может происходить в аэробных (в присутствии кислорода) или анаэробных (без кислорода) условиях.
Источник энергии: бактерии-фототрофы могут осуществлять фотосинтез, перерабатывая энергию солнечного света, а бактерии-хемотрофы используют в качестве источника энергии различные органические или неорганические соединения, осуществляя дыхание или брожение.
Природа окисляемого соединения. В соответствии с этим критерием выделяются органотрофы (для которых источником энергии служат органические соединения) и литотрофы, получающие энергию за счет окисления таких неорганических веществ, как H2, NH3, H2S, S, CO, Fe 2+ и т. д.
Источник углерода. Автотрофные бактерии фиксируют СО2, а гетеротрофные для получения углерода используют органические соединения.
Тип взаимоотношений с другими организмами. По типу взаимоотношений с другими организмамивыделяют сапрофитный (автономный) или паразитический (зависящий от организма-хозяина) образ жизни бактерий. Подверженность фаговой инфекции определенного типа также может служитьв качестве признака для классификации бактерий(phage typing).
Приспособленность к условиям среды обитания. В то время как мезофильные бактерии обитаютв умеренных условиях, другие бактерии (экстремофилы) приспособились к существованию в экстремальных условиях (температура, давление, рН, концентрация солей и т. д.). Признаки для дальнейшейклассификации бактерий можно обнаружить при изучении пигментации, анализе химических компонентов клеточной стенки и клеточной мембраны (составжирных кислот), данных иммунологического анализаповерхностных антигенов (серология) или устойчивости к действию антибиотиков. В последнее время особенно широко применяется анализ генетическихпризнаков. Первичные данные для классификацииможно получить из анализа состава ДНК (содержание G + C). Число бактерий, геном которых полностью секвенирован, постоянно увеличивается, и полученная информация используется при объяснениирезультатов анализа генетических признаков.С 1972 г. для классификации и установления эволюционных связей между различными бактериями используют результаты секвенирования рибосомныхРНК, прежде всего 16S- и 23S-рРНК. В этих молекулах выделяются высококонсервативные участки, сохранившиеся в процессе эволюции. В соответствиисо структурой рРНК все живые организмы делятся на три основных надцарства: археи, эубактерии (прокариоты) и эукариоты.
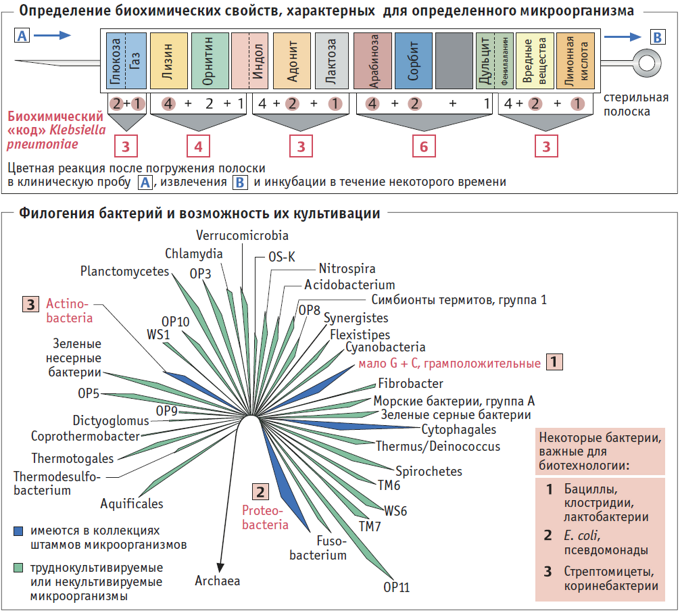
ХАРАКТЕРИСТИКА И ТАКСОНОМИЯ БАКТЕРИЙ. Быстрая и достоверная идентификация микроорганизмов имеет большое значение в клинической медицине, ветеринарной практике, пищевой промышленности и лабораторных исследованиях. Наряду с визуальным (с помощью микро скопа) и биохимическим анализом микроорганизма, изучают его способность расти на различных питательных средах, а также проводят анализ ДНК, например, используя ДНК-зонды, специфичные для определенных так со нов. Не всегда по полученным результатам можно отнести исследуемый штамм к тому или иному таксону, и в таких случаях требуется тщательный анализ более широкого набора признаков.

Некоторые бактерии, важные для биотехнологии
ВВЕДЕНИЕ. В качестве примеров бактерий, имеющих особенно важное биотехнологическое применение, мы рассмотрим следующие: Escherichia coli, Pseudomonas putida, Bacillus subtilis, Streptomyces coelicolor и Corynebacterium glutamicum.
ESCHERICHIA COLI – представитель кишечной флоры млекопитающих и принадлежит к группе энтеробактерий. Палочкообразные клетки имеют жгутики. E. coli – грамотрицательная бактерия, следовательно, под клеточной стенкой располагаются внешняя и внутренняя клеточные мембраны, разделенные периплазматическим пространством. В анаэробных условиях E. coli получает энергию в процессе брожения, а при наличии кислорода – с помощью дыхания. В оптимальных аэробных условиях продолжительность жизненного цикла (время между образованием клетки и ее делением) составляет около 20 мин. Геном E. coli имеет размер 4,6 млн п.н., и содержание GC-пар 51%. Нес мо тря на то что геном E. coli полностью секвенирован, и эта бактерия является одним из наиболее хорошо изученных микроорганизмов, в настоящее время известны функции лишь двух третей ее белков. В биотехнологии клетки E. coli используют для экспрессии негликозилированных белков, например инсулина, гормонов роста и фрагментов антител. Дикие штаммы E. coli относятся к «условно патогенным», так как обитают в кишечнике человека, поэтому в лабораторных экспериментах обычно используют ослабленные штаммы E. coli (например, E. coli К12), не представляющие угрозы для исследователей. Эти штаммы соответствуют группе безопасности S1, и их можно культивировать при соблюдении техники безопасности при обычных микробиологических экспериментах. Для клонирования чужеродной ДНК в E. coli используют различные векторы. В качестве примера мы выбрали ВАС-вектор, который наиболее часто применяют при создании генных библиотек (см. рисунок).
PSEUDOMONAS PUTIDA. Клетки P. putida – прямые палочки с полярными жгутиками. Это аэробные бактерии, обитающие в воде. Клетки не окрашиваются по Граму, т. е. под клеточной стенкой находятся две мембраны, а между ними – периплазматическое пространство. Размер генома P. putida составляет 6,1 млн п.н., содержание GC-пар – 61%. Бактерии P. putida имеют особенно важное значение при биотехнологической очистке окружающей среды, так как они способны разлагать трудноразлагающиеся вещества, в том числе ароматические соединения. Такое свойство обусловлено наличием в клетках бактерии так называемых плазмид деградации.
BACILLUS SUBTILIS (сенная палочка) – аэробная почвенная бактерия. Клетки имеют форму палочек без жгутиков. При неблагоприятных условиях в клетках B. subtilis формируются споры, устойчивые к изменениям температуры. B. subtilis относится к грамположительным бактериям, следовательно, под клеточной стенкой находится одна клеточная мембрана. Энергию бактерии получают в результате дыхания. В оптимальных условиях продолжительность жизненного цикла составляет около 20 мин. Геном B. subtilis имеет размер 4,2 млн п.н. и к настоящему времени он полностью секвенирован. Содержание оснований G + C составляет 44%. В биотехнологии штаммы B. Subtilis используют прежде всего для получения секретируемых ферментов, например протеаз и амилаз.
STREPTOMYCES COELICOLOR – почвенная бактерия, которая относится к группе актиномицетов. Все представители этой группы имеют хорошо развитый мицелий. На поверхности колоний образуется воздушный мицелий (гифы), а на концах гиф формируются споры (конидии). Актиномицеты относятся к грамположительным бактериям, т. е. под клеточной стенкой находится одна клеточная мембрана. Как и другие представители актиномицетов, S. coelicolor разрушает такие сложные органические соединения, как целлюлоза и хитин. Геном S. coelicolor почти вдвое больше, чем геном E. coli, – 8,7 млн п.н., и для него характерно высокое содержание оснований G + C (72%). Секвенирование генома S. coelicolor завершено. В результате выявлено почти 8000 структурных генов. Вероятно, такой большой геном содержит информацию для осуществления вторичного обмена веществ, например биосинтеза антибиотиков.
CORYNEBACTERIUM GLUTAMICUM принадлежит к группе коринебактерий. Представители этой группы обитают в самых разнообразных средах, и некоторые являются возбудителями болезней (например, C. diphteriae – возбудитель дифтерии). C. glutamicum – аэробные грамположительные бактерии. Клетки имеют булавовидную форму. Геном C. glutamicum размером 3,1 млн п.н. полностью секвенирован, содержание оснований G + C составляет 56%. Мутантные штаммы C. glutamicum являются важными продуцентами L-глутаминовой кислоты и L-лизина.
СЕКВЕНИРОВАНИЕ БАКТЕРИАЛЬНЫХ ГЕНОМОВ. К 2005 г. было завершено секвенирование геномов более200 бактерий. Среди них – многие патогенные бактерии, а также археи.


Грибы
ВВЕДЕНИЕ. Грибам принадлежит важная роль в круговороте веществ в природе, в частности они участвуют в разрушении древесины и образовании гумуса. В симбиозе с высшими растениями (микоризы) грибы помогают им перерабатывать различные вещества. В то же время некоторые представители грибов являются опасными патогенами растений (например, мучнистая роса). В биотехнологии грибы используют в производстве пищевых продуктов, при получении антибиотиков и ферментов; при утилизации биомассы важную роль играет способность грибов осуществлять разложение органических соединений. Среди 70 000 видов охарактеризованных грибов наиболее широко представлены сумчатые грибы аскомицеты (около 20 000 видов). В качестве примеров здесь рассмотрены Penicillium notatum и Aspergillus niger. Очень важны в биотехнологии представители фикомицетов – грибы родов Rhizopus и Mucor. Около 12 000 видов грибов базидиомицетов относятся к съедобным грибам (шампиньоны, лисички, белые грибы и т. д.), а некоторые другие их представители участвуют в разрушении древесины (белый и красный плесневые грибы). Среди грибов встречаются патогенные для человека виды (их около 300). Все грибы – гетеротрофы. Их клеточная стенка содержит хитин, глюкан и в некоторых случаях также целлюлозу.
РАЗМНОЖЕНИЕ ГРИБОВ. Типы размножения грибов чрезвычайно разнообразны, в качестве примера мы рассмотрим размножение сумчатых грибов аскомицетов. Вегетативное тело грибов (таллом) состоит из мицелия – системы ветвящихся нитей (гифы). При бесполом размножении на концевых выростах мицелия образуются конидиеносцы. В конидиях происходит деление и высвобождение спор, из которых формируется новый мицелий. Как и многие другие грибы, аскомицеты способны к половому размножению. У них существует две различающиеся по облику формы (явление диморфизма), что существенно затрудняет их классификацию. Гифы формируют мужские и женские половые органы (антеридии и аскогонии соответственно); после протекания плазмогамии образуются дикариотические гифы, дающие начало аскокарпу («плодовому телу»). Затем в концевых клетках дикариотических гиф два ядра сливаются и формируют диплоидную зиготу (этот процесс называется кариогамией). После мейоза образуются восемь аскоспор (у аскомицетов) или четыре базидиоспоры (у базидиомицетов), из которых в дальнейшем вырастает мицелий.
PENICILLUM NOTATUM растет как мицелий. Он образует плодовые тела с конидиями, из которых высвобождаются споры. Внутри асков созревают споры, прорастающие в новый мицелий. Как и другие представители несовершенных грибов, P. notatum размножается исключительно бесполым путем, поэтому для осуществления рекомбинации в лабораторных условиях проводят слияние протопластов с различными типами ядер (гетерокариоз, парасексуальный процесс). P. notatum и родственный гриб Cephalosporium acremonium – продуценты важнейших лактамных антибиотиков. Другим примером биотехнологического применения грибов рода Penicillium является использование Penicillium camamberti при изготовлении сыров. Геном P. notatum имеет размер 32 млн п.н., и к настоящему времени секвенирована лишь небольшая его часть.
ASPERGILLUS NIDULANS – другой представитель аскомицетов, морфологически отличающийся от Penicillium по форме конидий. Размер генома составляет 31 млн п.н. и его структура пока полностью не расшифрована (2005 г.). В биотехнологии штаммы Aspergillus находят широкое применение: A. oryzae часто служит в качестве организма-хозяина для экспрессии рекомбинантных белков, A. niger является промышленным продуцентом лимонной и глюконовой кислот. Другие штаммы Aspergillus используются для синтеза внеклеточных ферментов (амилаз и протеиназ), а также в пищевой промышленности (например, в азиатском регионе для приготовления соевого соуса и мисо). Как и в случае Penicillium, для усовершенствования штаммов Aspergillus проводят слияние протопластов с последующим отбором.
RHIZOPUS ORYZAE И R. NIGRICANS – представители фикомицетов, растущие на рисе и хлебе соответственно. Гифы этих грибов растут чрезвычайно быстро и буквально пронизывают питательную среду. Бесполое размножение фикомицетов происходит с образованием спор в специализированном мицелии – спорангиях. Представители родов Rhizopus и Mucor находят широкое применение в биотехнологии, так как в процессе разложения органического субстрата они выделяют в среду разнообразные внеклеточные ферменты. Геном R. oryzae составляет около 39 млн п.н., его нуклеотидная последовательность будет определена в ближайшее время (2005 г.).


Дрожжи
ВВЕДЕНИЕ. Дрожжи – это группа грибов, не имеющих типичного мицелия и размножающихся почкованием. К дрожжам относят представителей различных классов грибов. Дрожжи – гетеротрофные организмы, предпочитающие для роста кислые среды (рН 3,5–5,0). Клеточная стенка этих организмов содержит хитин. В биотехнологии наиболее важные следующие дрожжи: Saccharomyces cerevisiae, Candida utilis, Candida albicans, Schizosaccharomyces pombe, Hansenula polymorpha и Pichia pastoris.
SACCHAROMYCES CEREVISIAE (пекарские дрожжи) могут размножаться как в гаплоидной, так и в диплоидной форме, поэтому они являются излюбленным объектом для генетических исследований. Гаплоидные штаммы, используемые в лабораторной практике, принадлежат одному из двух типов спаривания – MATa или MATα. Скрещивание возможно только между представителями разных типов спаривания. Бесполое (вегетативное) размножение осуществляется почкованием гаплоидных или диплоидных клеток. При половом размножении происходит слияние двух гаплоидных клеток, и в результате мейоза образуются четыре гаплоидные аскоспоры. Выводы о генетических изменениях можно делать, исходя из визуального (с помощью микроскопа) анализа аскоспор (тетрадный анализ). Поскольку и в гаплоидной, и в диплоидной формах эти дрожжи – неприхотливая и устойчивая культура, для которой в оптимальных условиях время удвоения составляет всего 90 мин, S. cerevisiae – очень ценный объект для молекулярно-генетических экспериментов. Нуклеотидная последовательность генома S. cerevisiae полностью расшифрована (12 млн п.н. находятся в 46 хромосомах). Наличие в клетках внехромосомных элементов (2 -плазмиды, присутствующей в количестве 60–100 копий на клетку) также является преимуществом этого вида дрожжей, так как открывает новые возможности для осуществления рекомбинации. Для трансформации клеток S. cerevisiae разработано множество векторных конструкций: при использовании одних векторов чужеродная ДНК реплицируется в клетке независимо (YRP – yeast replicating plasmid и YEP – yeast episomal plasmid), а другие век торы позво ляют чужеродной ДНК встраи ваться в дрожжевые хромосомы (YIP – yeast integrating plasmid). Искусственные дрожжевые хромосомы (YAC – yeast artificial chromosome) позволяют клонировать очень крупные фрагменты ДНК (600–1400 т. п. н.), поэтому их часто используют для создания геномных библиотек. Недостатком такой системы является высокая частота внутри хромосомной рекомбинации, поэтому в последнее время искусственные дрожжевые хромосомы все чаще заменяют на бактериальные искусственные хромосомы (ВАС). Около 6000 генов S. cerevisiae в высокой степени гомологичны генам человека, поэтому эти дрожжи часто служат в качестве модельной системы при изучении молекулярно-биологических механизмов обмена веществ. В биотехнологии они используются в пищевой промышленности, для производства алкогольных напитков, этанола, а также для промышленного получения рекомбинантных белков, например α-интерферона, и различных вакцин, в том числе поверхностного антигена вируса гепатита В. В отличие от систем экспрессии с использованием E. coli, в клетках S. cerevisiae белки претерпевают посттрансляционную модификацию (например, гликозилирование), поэтому дрожжи используют для получения эукариотических белков, для функционирования которых необходимы такие модификации.
CANDIDA UTILIS – представитель группы дрожжей, который формирует мицелий и размножается исключительно почкованием. В геноме C. utilis встречаются неканонические кодоны (например, CUG, обычно кодирующий лейцин, в этом организме кодирует серин), поэтому гетерологическая экспрессия в C. utilis крайне затруднена. Штаммы C. utilis применяют в биотехнологическом производстве внеклеточных ферментов, а в сельском хозяйстве – как кормовую добавку. Грибы рода Candida могут использовать в качестве источника углерода такие необычные вещества, как фракции нефти или сульфитсодержащие производные, поэтому они имеют особенно важное значение при утилизации различных отходов. Некоторые представители рода Candida для человека патогенные (Candida albicans).
PICHIAPASTORIS И HANSENULA POLYMORPHA – представители метилотрофных дрожжей, использующих в качестве единственного источника углерода метанол. Метилотрофные дрожжи активно изучаются с целью использования для экспрессии эукариотических генов. Так, в клетках Pichia pastoris удалось осуществить очень эффективный синтез различных белков: липаз, β-интерферона и фрагментов антител (до 12 г рекомбинантного белка на литр культуры).
SCHIZOSACCHAROMYCES POMBE впервые были выделены из восточно-африканского пива (пиво на язык суахили – pombe). Геном S. pombe, распределенный всего между тремя хромосомами, полностью расшифрован: его размер составляет 12,6 млн п.н., что сравнимо с размером генома S. cerevisiae, но всего по трем хромосомам, несущим почти по 5000 генов.



Микроорганизмы: выделение и хранение штамма. Техника безопасности
ВВЕДЕНИЕ. В научных исследованиях, как правило, используют чистые культуры микроорганизмов. Для биотехнологического применения штаммы усовершенствуют в соответствии с поставленными задачами, осуществляя мутагенез и последующий отбор мутантов. Для сохранения чистых штаммов микроорганизмов создают специальные коллекции. Пересев клеток микроорганизмов на жидкие или твердые питательные среды проводится в стерильных условиях. Большинство используемых в биотехнологии микроорганизмов являются гетеротрофами и культивируются в аэробных условиях. Анаэробные организмы выращивают в бескислородной среде, а для культивирования фотосинтезирующих микроорганизмов подбирают специальные условия освещенности.
ЧИСТЫЕ МИКРОБНЫЕ КУЛЬТУРЫ хранятся в коллекциях штаммов. Чтобы получить чистую культуру из природной среды обитания (почвы, воды, пищевых продуктов, других организмов), проводят посев штрихом (метод истощающего штриха) на стерильную агаризованную питательную среду (агар – сложный полисахарид, получаемый из морских водорослей). Условия культивирования выбирают так, чтобы выделяемый микроорганизм имел преимущество роста по сравнению с другими микроорганизмами, обитающими в той же природной среде. Например, цианобактерии культивируют в анаэробных условиях на свету, в качестве источника углерода пропускают СО2, а в качестве источника азота – N2. Для выращивания грибов используют слабокислую, обогащенную сахарами среду, для отбора термофильных микроорганизмов повышают температуру культивирования, а для выделения микроорганизмов продуцентов протеаз в качестве единственного органического источника азота в среду добавляют казеин. По данным 16S-рРНК-анализа удается выделить лишь 1% всех микроорганизмов, обитающих в воде или почве.
КОЛЛЕКЦИИ МИКРООРГАНИЗМОВ служат для хранения чистых штаммов, обладающих определенными свойствами. Для поддержания штамма в лабораторных условиях его пересевают на агаризованную среду: в пробирку со скошенной средой или на чашку Петри, однако в результате многократного пересева может произойти вырождение. По этой причине для сохранения особенно важных штаммов предпочтительнее использовать один из следующих методов: 1) хранение под химически инертной жидкостью, на- пример парафином (рекомендуется для сохранения штаммов гифообразующих грибов); 2) замораживание при –196°С и последующее хранение в жидком азоте или при температуре –70 °С. Замораживание и размораживание в этом случае должны происходить очень быстро и в присутствии глицерина, чтобы предотвратить разрушение клеточной стенки образующимися кристаллами льда. Метод широко применяется для сохранения бактериальных и дрожжевых штаммов; 3) высушивание клеточной суспензии на носителе (силикагеле, песке) в вакууме. Эту операцию проводят в присутствии эмульгаторов (обезжиренное молоко, сыворотка), а полученные препараты хранят при температуре –70 °С. Перед помещением в коллекцию необходимо убедиться в том, что клетки не потеряли способности к росту. В настоящее время в большинстве стран созданы богатые коллекции штаммов микроорганизмов, из которых можно получать чистые культуры. Существуют как универсальные коллекции штаммов, например ATCC – американская коллекция штаммов (American Type Culture Collection) или DSMZ – коллекция микроорганизмов и клеток Германии, так и коллекции, специализирующиеся на определенных группах микроорганизмов, например CBS – центральная коллекция плесневых грибов. Многие промышленные предприятия имеют собственные коллекции важных штаммов микроорганизмов.
ТЕХНИКА БЕЗОПАСНОСТИ. Почти в любом роде микроорганизмов есть представители, патогенные для человека: например, в биотехнологии Bacillus subtilis – непатогенный продуцент ферментов, а вот Bacillus anthracis – возбудитель сибирской язвы; Aspergillus oryzae используется для приготовления соевого соуса, а Aspergillus flavus продуцирует токсичное вещество (афлатоксин), обладающее канцерогенными свойствами. По этой причине при любых операциях с микроорганизмами необходимо строго соблюдать правила техники безопасности при работе с биологическими объектами. В соответствии со степенью риска для пользователя все микроорганизмы разделены на четыре группы. Оборудование лаборатории и правила работы должны соответствовать правилам техники безопасности при работе с микроорганизмами каждой группы. К первой группе риска относятся микроорганизмы, которые традиционно используются для приготовления пищевых продуктов (например, пекарские дрожжи). Подавляющее большинство микроорганизмов, использующихся в биотехнологии, также относятся к первой группе.


Усовершенствование штаммов микроорганизмов
ВВЕДЕНИЕ. Микроорганизмы, выделенные из природной среды обитания, редко полностью отвечают требованиям биотехнологического производства. Поэтому необходимым подготовительным этапом является многократное повторение мутагенеза и последующего отбора мутантов, обладающих искомыми свойствами. Наиболее часто задачей таких экспериментов является получение штаммов, продуцирующих больше целевого вещества и меньше побочных продуктов или обладающих лучшими технологическими характеристиками (сокращение времени ферментации, отсутствие нежелательной пигментации, устойчивость к бактериофагам и др.). Одним из преимуществ микроорганизмов является их короткий жизненный цикл (часто менее 1 ч). Это позволяет достаточно быстро получать и анализировать большое количество мутантов. При работе с эукариотическими микроорганизмами (например, грибами) необходимо принимать во внимание процесс рекомбинации. Знание обмена веществ, его регуляции и структуры ДНК, кодирующей ферменты, позволяет модифицировать геном микроорганизмов, целенаправленно «выключая» или «активируя» те или иные стадии обмена веществ (metabolic engineering).
МУТАГЕНЕЗ. Вероятность спонтанной мутации в нормальном гене (под действием естественных мутагенов или при репликации) составляет 10–6–10–7. Как правило, такие мутации остаются «молчащими» из-за генетической или функциональной супрессии или становятся мишенью системы репарации ДНК. Для усовершенствования штаммов осуществляют более эффективный искусственный мутагенез: например, при ультрафиолетовом облучении или при введении мутагенных веществ. Путем изменения длительности мутагенного воздействия и путем подбора мутагенных веществ удается варьировать частоту мутаций в клетках. В результате действия мутагенных факторов, как правило, 90–99% клеток погибают, а затем из оставшихся клеток отбирают мутантов, которые обладают желаемыми фенотипическими признаками.
ОТБОР МУТАНТОВ В ПОВЕРХНОСТНЫХ КУЛЬТУРАХ.
Основным фенотипическим признаком, рассматриваемым при усовершенствовании штаммов микроорганизмов, является их способность образовывать большое количество продукта. Для отбора мутантов, удовлетворяющих этому требованию, разрабатывают систему отбора. В качестве признака для отбора может служить устойчивость к антибиотику, ядовитым веществам или фаговой инфекции. Полученные после проведения мутагенеза клетки высевают в чашки Петри на твердые селективные среды, где вырастают лишь те колонии, которые обладают необходимой устойчивостью. Для отбора ауксотрофных мутантов реплики колоний помещают на питательную среду без антибиотика и с антибиотиком (например, с пенициллином). Основными преимуществами такого метода является возможность проводить одновременный анализ большого количества полученных мутантов (несколько сотен на одной чашке). В результате действия сильных мутагенных факторов часто происходят изменения в нескольких генах, поэтому необходимо проверить не только способность мутантов образовывать необходимый продукт, но и сохранность свойств, характерных для дикого штамма. Для этого отобранные клетки выращивают в качалочных колбах в условиях, максимально приближенных к таковым в производственном процессе, и отбирают клетки, обладающие улучшенными свойствами. Повторение нескольких раундов мутагенеза с последующим отбором позволяет отобрать штаммы, которые наилучшим образом удовлетворяют всем требованиям технологического процесса. Для устранения ненужных приобретенных свойств полученные штаммы скрещивают с дикими штаммами.
СЕЛЕКЦИЯ В НЕПРЕРЫВНОЙ КУЛЬТУРЕ. Отбор мутантов можно проводить в непрерывной культуре в ферментере: для этого клетки микроорганизмов выращивают в среде с мутагеном и осуществляют при этом селективное воздействие, например постепенно заменяют один источник углерода на другой. В таких условиях выживают лишь те микроорганизмы, которые в результате мутагенеза приобрели способность утилизировать новый источник углерода. Описанный способ, однако, не позволяет проводить отбор мутантов с повышенным выходом продукта.
Болезни животных, которые могут передаться человеку

Домашние питомцы приносят радость в дом. Общение с ними сближает человека с природой, делает его добрее, дарит тепло и заботу. Для многих собачка, кошечка или хомячок являются полноценными членами семьи, с которыми, как с детьми, проводится большая часть времени. Бывая в зоопарках, малыши и взрослые пытаются покормить, приласкать самых разнообразных животных, при этом забывая, что они, как и люди, могут болеть или являться носителями вирусных или инфекционных заболеваний различной степени тяжести. Употребляя в пищу некачественное мясо, молоко, яйца, также можно перенести заболевание в свой организм.
Болезни, передающиеся через животных
Бактерии, вирусы и различного рода инфекции могут попасть в организм человека несколькими путями:

- Через непосредственный контакт с животными. Обычные ласки, поглаживания, поцелуи могут надолго испортить настроение и здоровье детям и взрослым. Также одним из источников служат грязные лапы, шерсть. Пройдя босыми ногами по грязному полу, садясь на диван или ложась в постель, на которой до этого побывало грязное животное, человек может стать жертвой кожных заболеваний.
- Через продукты питания. Используя продукты животноводства, не прошедшие определенную обработку, можно быстро приобрести вирусную или бактериальную инфекцию.
- Через воду. Даже купание в реке или употребление в пищу воды из ручья или пруда, находящихся рядом с пастбищами, может стать источником заболеваний.
Болезни животных, передающиеся человеку при непосредственном контакте
Эта группа заболеваний наиболее распространена. Прогулки и игры с животными на природе, кормление их в зоопарке или на водоеме – безобидные факторы приобретения заболеваний.

- Если вы получили легкую царапину или укус от зараженной кошки, или она лизала любую ранку на теле – есть вероятность получения инфекционного заболевания, вызванного определенными видами бактерий. Важно, что кошка при этом не испытывает дискомфорта, а служит лишь переносчиком заболевания. У человека инкубационный период может длиться до двух недель и в дальнейшем сопровождаться повышением температуры, воспалением лимфатических узлов и т.д. Заболевание неопасно для человека, но если не заниматься его лечением, может принести негативные последствия. Это, так называемая болезнь, кошачьих царапин.
- При укусе зараженной собакой может развиться вирусное заболевание, именуемое бешенством. Его симптомы: жар, слабость, тревожность, головная боль. В этом случае лечение должно быть незамедлительным и сопровождаться вакцинацией.
- Если морская свинка или хомячок — носители лимфоцитарного хориоменингита, болезнь может передаться человеку через укус, слюну и т.д. Симптоматика заболевания аналогична с гриппом: повышение температуры, жар, слабость, ломота и т.д. В зависимости от тяжести заболевания может потребоваться стационарное лечение.
- Птицы, особенно попугаи, могут стать переносчиками орнитоза. Заболевание проникает в организм при чистке клетки, через глазные выделения, когда птица клюет вас или получает пищу изо рта. В этом случае оно проявляется в виде признаков, сходных с пневмонией. Лекарственными средствами служат антибиотики.
- Кожное грибковое заболевание – стригущий лишай – передается при прикосновении к больному участку животного. На коже человека он проявляется в виде пятен различных размеров, которые вызывают зуд и сухость кожи. При отсутствии лечения ореол пятна может расширяться. Лекарственными препаратами при лечении лишая являются антигрибковые мази и гели.
Болезни животных, передающие через мясо и молоко
Промышленное производство мяса, птицы и крупного рогатого скота, свиней, овец и т.д. подразумевает строгий контроль по всем видам показателей, в том числе бактериальных. Но если приобретаются продукты у неизвестного производителя, есть вероятность их заражения.

- При употреблении зараженного бруцеллезом молока, сыра и других видов молочной продукции есть риск заразиться этим бактериальным заболеванием. Озноб, дрожь, высокая температура, утомляемость и мышечные боли – основные симптомы болезни. Лечение подразумевает прием антибиотиков.
- Сальмонелла может вызвать одноименное заболевание «сальмонеллез». Употребляя в пищу зараженное мясо или яйцо птицы (кур, гусей), можно заразиться сальмонеллой. Распространяясь через кровь, она оказывает разрушающее действие на печень, почки и другие органы.
Меры профилактики
Чтобы избежать негативной стороны контакта с животными, соблюдайте простейшие меры предосторожности: содержите домашних питомцев в чистоте, регулярно проводите вакцинации, мойте руки не только перед едой, но и после контакта с животными. Особое внимание уделяйте маленьким детям, которые через рот могут внести любую инфекцию.
Мясо следует подвергать длительной варке: при жарке, копчении и других похожих видах обработки источники заболевания могут сохраниться. Молоко следует хорошо пастеризовать.
Относитесь к своему здоровью и здоровью близких внимательно, соблюдайте условия гигиены и простейшие правила безопасности, следите за домашними питомцами и будьте здоровы.
http://propionix.ru/osnovy-mikrobiologii
https://www.snta.ru/press-center/bolezni-zhivotnykh-kotorye-mogut-peredatsya-cheloveku/
