Bio-Lessons
Образовательный сайт по биологии
Вирусы. Бактериофаги. Вирусные заболевания.
Вирусы. Бактериофаги. Вирусные заболевания.
Вирусы (от лат. вирус — яд) – представляют собой мельчайшие неклеточные формы жизни. Вирусы имеют размеры 2-5*10 -7 см, что значительно меньше, чем бактериальная клетка (от 0,2 до 10 мкм). Рассмотреть вирусы возможно только с помощью электронного микроскопа, увеличивающий в 100 тысяч и более раз. Вирусы относятся к отдельному царству.
Вирусология – наука изучающая вирусы. Становление вирусологии как науки начинается с 30-х годов 20 века.
История открытия вирусов
Впервые вирус табачной мозаики (рис.1) был открыт русским ученым Д.И.Ивановским (1892г.) (рис.2).

Рис.1 Листья табака (слева) пораженные вирусом табачной мозаики (справа)
Студент Петербургского университета Дмитрий Ивановский выезжал на Украину и в Бессарабию для определения причин болезни табака. В листьях табака будущий ученый не обнаружил клеток бактерий, однако было замечено, что сок зараженного растения поражал здоровые листья. Используя свечу Шамберлана Ивановский профильтровал сок больного растения, тем самым исключив прохождение через фильтр мелких бактерий. Полученный фильтрат все равно вызывал заражение листьев табака. Это еще раз доказывало «невиновность» бактерий. Д.И.Ивановский попробовал культивировать возбудителя на питательной среде, однако это не дало результата. После проведенных опытов Дмитрий Иосифович пришел к выводу, что возбудитель является необычной природы и имеет размеры в разы меньше чем клетка бактерии. В последствие возбудитель был назван «фильтрующиеся бактерии».
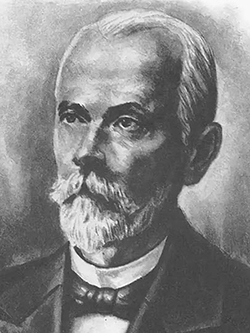
Рис.2 Д.И.Ивановский — первооткрыватель вирусов
Свои выводы ученый изложил в труде «О двух болезнях табака» в 1892 году. Именно этот год считается годом открытия вирусов.
Наряду с Д.И.Ивановским изучением вирусов занимался голландский микробиолог Мартин Бейеринк, который в 1898 году повторив опыты русского ученого, назвал вирусный раствор – «заразной живой жидкостью» или «жидкий живой контагий».
Первый вирус животных (вирус ящура) был описан в 1897 году Лёффером и Фрошем. В 1901 году вирус желтой лихорадки был открыт англичанами У. Ридом и Д. Кэрроллом.
В 1917 году Ф.д’Эррелем был открыт бактериофаг – вирус, поражающий бактерии.
Удивительно то, что первая вакцина от оспы была предложена за 100 лет до открытия вирусов, в 1796 году английским врачом Э.Дженнером. Второй по открытию стала – антирабическая вакцина, представленная французским ученым микробиологом Л.Пастером в 1885 году.
Названия «ультравирус» и «фильтрующий вирус» использовались в науке до укоренившегося ныне краткого термина — «вирус», который впервые применил Л.Пастер.
Строение и формы вирусов
Вирусы — неклеточные частицы, состоящие из белковой оболочки (капсид) и собственного генетического материала в виде нуклеиновой кислоты (ДНК или РНК) (рис.3).
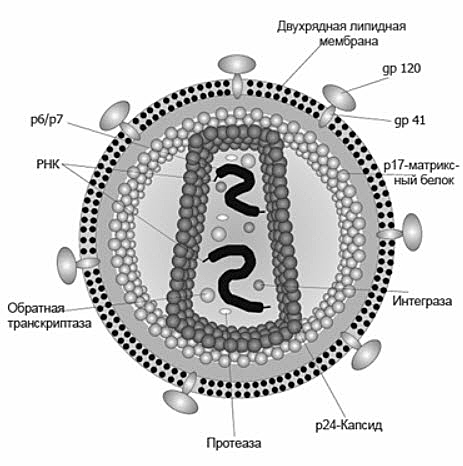
Рис.3 Строение вируса иммунодефицита человека (ВИЧ)
Вирусы могут обладать разнообразными формами: шаровидные, овальные, палочковидные, нитевидные, цилиндры, тетраэдры, октаэдры и др.(рис.4).
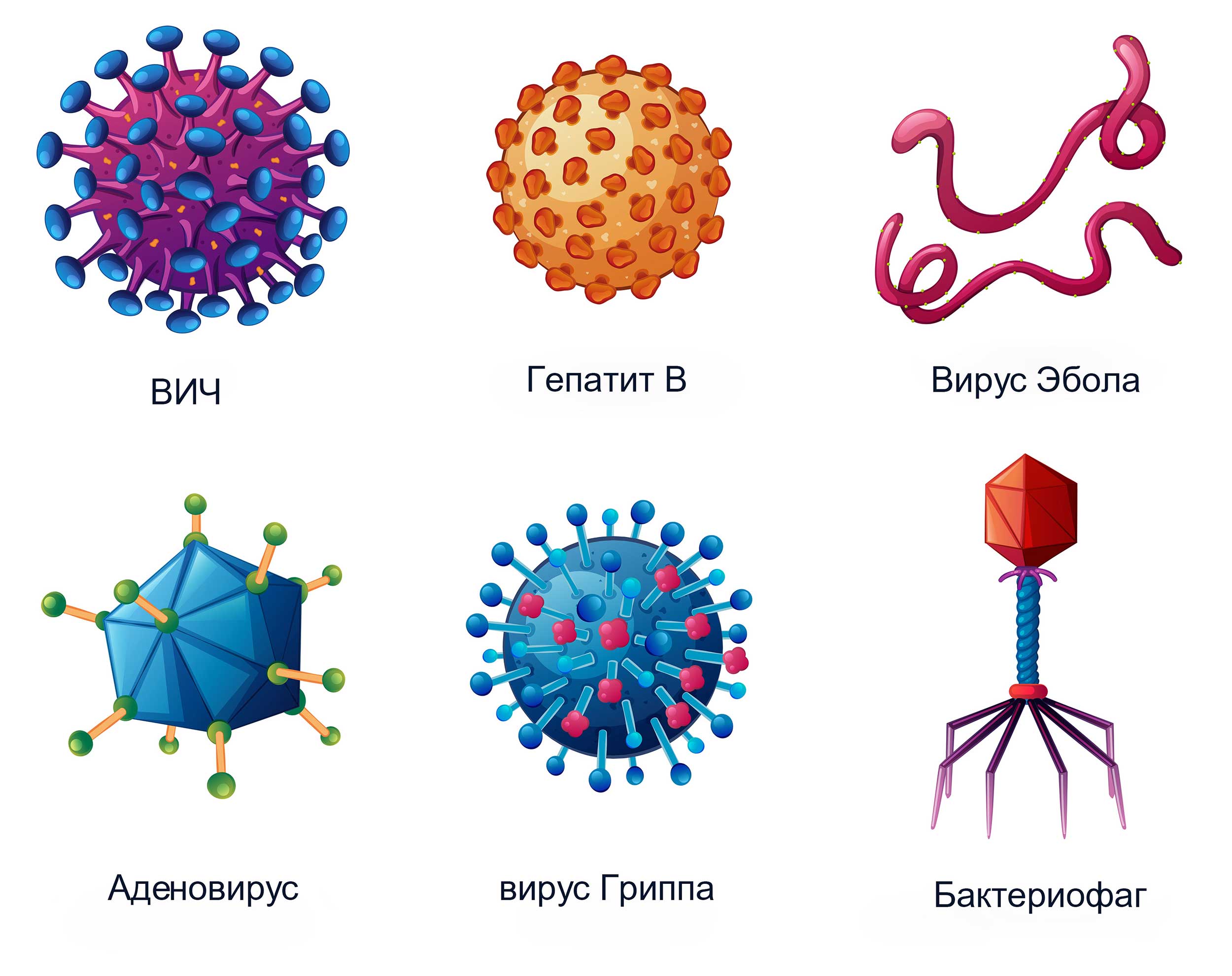
Рис.4 Разнообразные формы и виды вирусов
Более сложные вирусы имеют в своем составе дополнительные белковые или липопротеидные оболочки. Вирусы гриппа и герпеса кроме белковой оболочки могут содержать и углеводы.
| ДНК-содержащие вирусы | РНК-содержащие вирусы |
|---|---|
| оспы | бешенства |
| герпеса | кори |
| бактериофаги Т-группы | СПИДа и лейкоза |
| гепатита В | гепатита А |
| паповавирусы | гриппа |
| аденовирусы | полимиелита |
| цитомегаловирус | ОРЗ |
| Эпштейн-Бара | желтой лихорадки |
| и др. | краснухи и др. |
Геном вирусов может быть представлен однонитчатыми и двунитчатыми молекулами ДНК (вирус оспы человека, овец, свиней, аденовирус человека) и РНК (матрица для вирусов насекомых и других животных). Вирусы с однонитчатой молекулой РНК (энцефалит, краснуха, корь, бешенство, грипп и др.).
Вне живой клетки вирус не питается, не передвигается, не растет, не размножается и не проявляет других свойств живого.
Размножение вирусов
Вирусы способны размножаться только внутри живой клетки. Вирус проникает внутрь клетки путем связывания его с особым протеином – рецептором, расположенным на поверхности клетки. На поверхности чувствительной клетки происходит связывание с рецептором, после чего присоединившейся участок погружается в цитоплазму и превращается в вакуоль. Стенки вакуоли, состоящей из цитоплазматической мембраны способны сливаться с другими вакуолями или даже ядром. В результате вирус достигает любой участок клетки.
Основные этапы заражения вирусом:
1.Присоединение вируса к мембране клетки-хозяина.
2.Впрыскивание своей нуклеиновой кислоты (НК) в цитоплазму клетки-хозяина. Капсид остается снаружи.
3. ДНК вируса у эукариотической клетки проникает в ядро клетки и встраивается в ДНК хозяина. В бактериальной клетке ДНК вируса встраивается в ДНК бактерии. РНК содержащие вирусы вначале делают из нее копию в виде ДНК, а затем полученную ДНК встраивают в хромосому или нуклеотид клетки-хозяина.
4. Генетический материал клетки хозяина перестает функционировать. В ядре осуществляется синтез копий вирусной НК, а в цитоплазме на рибосомах – синтез копий вирусного капсида.
5. Образуются функциональные вирусы. НК «одевается» в капсид, происходит сборка вирусных частиц.
6. Образовавшиеся вирусы выходят из клетки-хозяина, так как первая исчерпала свои ресурсы. Вирусы продолжают проникать в новые клетки, расположенные поблизости.
Описанные этапы характерны для литических (от греч. «лизис» — растворение, разрушение, распад) вирусов, вызывающие разрушение или гибель клетки. Существует так же второй вид, так называемых «умеренных» вирусов. Они встраиваю свою нуклеиновую кислоту в ДНК клетки.
Паразитируя на генетическом уровне живой клетки, они встраиваются в ее геном. Внедрив свой генетический код в молекулу ДНК, вирус становится частью живой клетки. В такой форме он может не проявлять себя неопределенно долгое время. В какой-то момент вирусные частицы ДНК «включаются» одновременно во всех зараженных клетках, вызывая их гибель.
Данный процесс до конца не изучен, и возможно именно он мог бы решить вопрос возникновения онкологических заболеваний.
Быстрая способность адаптироваться и видоизменяться, подстраиваясь к геному клетки, делает некоторые вирусные заболевания практически неизлечимыми. Таким образом, вирусы представляют паразитизм на генетическом уровне (рис.5).
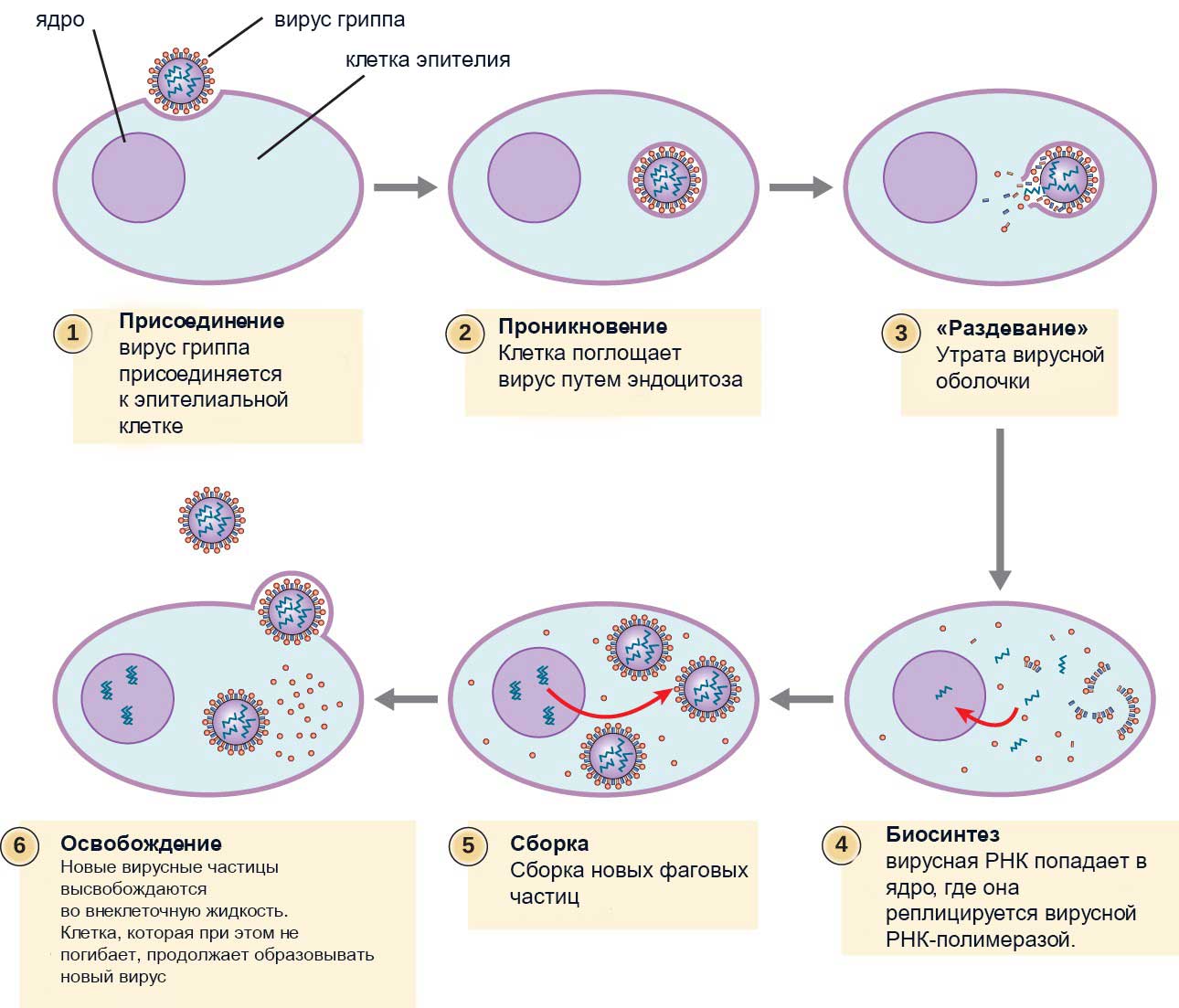
Рис.5 Размножение вируса гриппа
Совсем по-другому проникает в клетку вирус бактерий – бактериофаг (от греч. фагос – «пожирающий»)(рис.6).
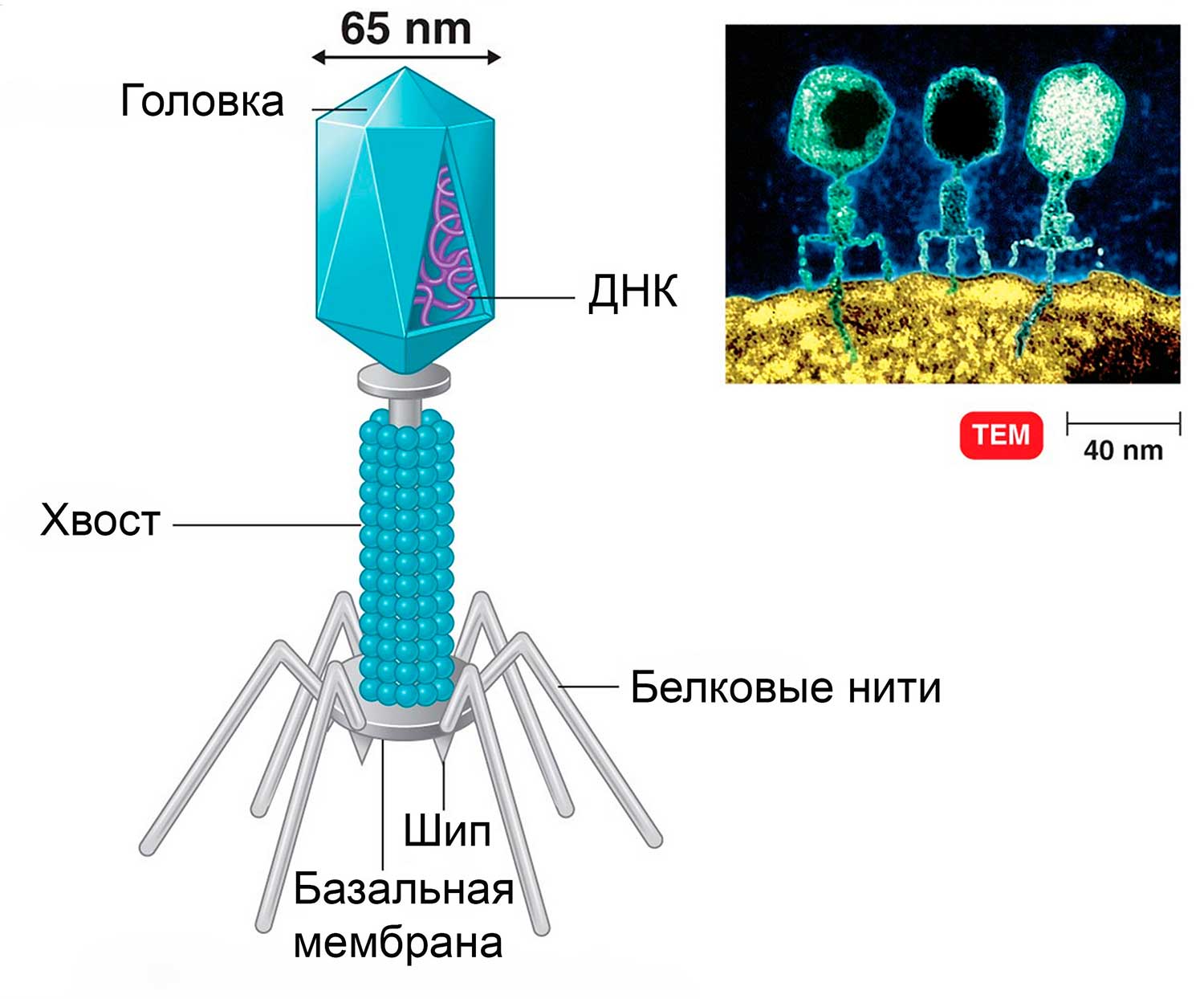
Рис.6 Строение бактериофага
Бактериофаг состоит из головки, хвостика и нескольких хвостовых отростков (белковых нитей). Наружная часть головки покрыта белковой оболочкой. Во внутренней части головки расположена ДНК, а внутри хвоста проходит центральный канал. Из-за толстых клеточных стенок бактерий белок-рецептор бактериофага не может погрузиться в цитоплазму.
Удерживаясь на поверхности клетки за счет шипов, расположенных под базальной мембраной, бактериофаг пронзает стенку бактерии и вводит внутрь полый стержень. По этому стержню в цитоплазму поступает ДНК (или РНК). Геном бактериофага проникает внутрь клетки, а оболочка остается снаружи. Спустя время, сформировавшиеся зрелые фаговые частицы разрушают бактерию изнутри и выходят наружу (рис.7).
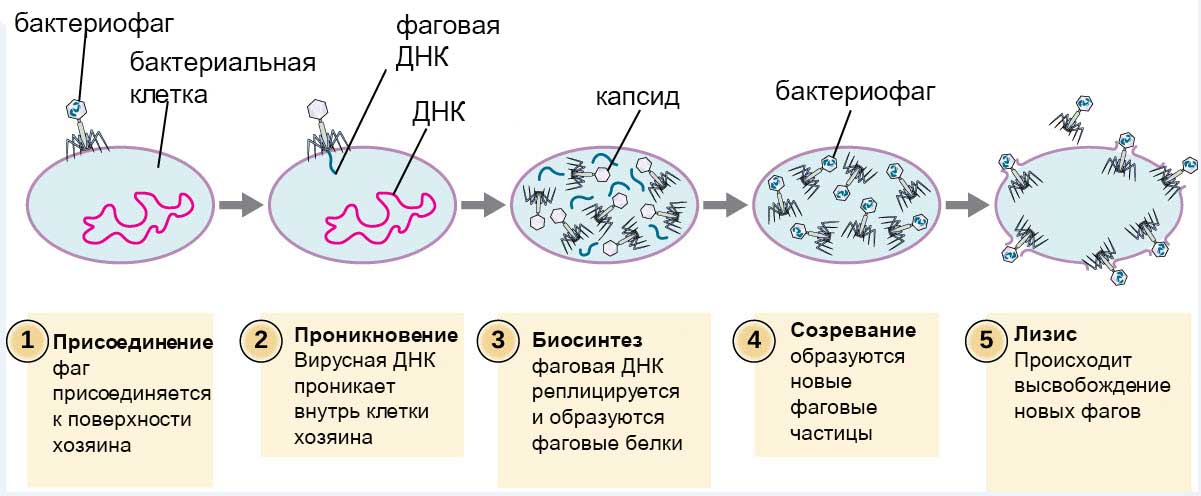
Рис.7 Размножение бактериофага
Обладая способностью полного уничтожения бактериальной клетки, бактериофаги могут быть использованы для лечения разнообразных бактериальных заболеваний (холеры, дизентерии, брюшного тифа и др.).
Отмечено, что отделение от вирусной частицы нуклеиновой кислоты приводит к потере инфекционной способности к репродукции. Это говорит о том, что нуклеиновая кислота играет важную роль в размножении вируса.
При благоприятных условиях вирус очень быстро размножается. Так, за 30 минут в одной клетке появляются сотни новых вирусов.
Вирусы могут продолжительно сохраняться в почве, воде и другим средах. Некоторые представители устойчивы к высоким температурам (свыше +100С) и высушиванию.
Виды вирусных заболеваний
В настоящее время известно около 400 видов вирусов растений и около 500 видов вирусов животных.Вирусы растений вызывают поражение листьев и других органов, вызывая появление разноцветных или бесцветных пятен и полосок. Вирусы вызывают замедление роста растений, изменяет их форму и снижает урожайность.
Наиболее опасными для человека являются вирусы гепатита – А, В, С. Вирус способен сильно повреждать ткани печени, вызвав необратимые последствия.
ВИЧ.СПИД
Более опасную для человечества форму представляет вирус иммунодефицита человека или сокращенно ВИЧ (HIV). Попав в кровь, ВИЧ, поражает иммунную систему человека, приводя к развитию болезни под названием СПИД (синдром приобретенного иммунодефицита человека). РНК-содержащий ВИЧ атакует белые кровяные клетки – лимфоциты, отвечающие за иммунитет, делая человека уязвимым для других болезней.
Внедренный в лимфоциты РНК вирус начинает синтезировать фермент – ревертазу. Этот фермент служит матрицей для последующего синтеза молекулы ДНК. Синтезированная вирусная ДНК встраивается в хромосому лимфоцита. После чего вирус долгое время может не проявлять себя. Это может длиться от 1 до 2 лет, а иногда и более. Спустя время вирусная ДНК начинает проявлять себя, синтезируя сотни тысяч вирусов, что в итоге приводит к разрушению лимфоцита.
Вероятность заражения ВИЧ увеличивается при прямом контакте с кровью больного человека. Распространенные пути передачи вируса; незащищенный половой контакт с инфицированным человеком, инъекции шприцом, переливание крови. ВИЧ не передается воздушно-капельным путем, через укусы насекомых, посуду, при рукопожатиях и пользовании общественными местами (туалеты, бассейны, бани и т.п.).
В настоящее время вакцины против СПИДа нет, но существуют медицинские препараты на основе азотимидина и ингибиторов протеаз, способные подавить синтез вирусной ДНК. Это облегчает течение болезни и значительно удлиняет жизнь человека.
Хочется отметить, что ВИЧ инфицированный человек, вовремя обратившийся в центр СПИДа, может контролировать развитие этой тяжелой болезни и в принципе жить полноценной жизнью с соблюдением определенных мер. При отсутствии строгого контроля и лечения, стадия инфицирования переходит в стадию СПИДа, которая неминуемо ведет к гибели. Человек на стадии СПИДа, из-за «иммунной беспомощности», может погибнуть от ряда инфекционных болезней.
Симптомами СПИДа является температура, постоянный озноб, легкая простужаемость, резкое похудение.
Чтобы предупредить СПИД необходимо соблюдать следующие правила;
— избегать прямого контакта с кровью неизвестного человека (зараженными так же могут быть лимфа, сперма, влагалищные выделения, грудное молоко и др.);
— избегать случайные половые связи;
— использовать презервативы;
— пользоваться одноразовыми шприцами;
— пользоваться личными бритвенными приборами, при этом не разрешать пользоваться своими.
Природным очагом СПИДа по мнению ученых считается Центральная Африка, а носителем вируса являются зеленые мартышки.
Грипп
Всем известный вирус гриппа не менее опасный, наряду с корью, гепатитом и полиомиелитом.
Грипп – болезнь, угрожающая человеческой жизни. В 1918-1919 годах весь земной шар трижды был охвачен волнами гриппа, во время которых погибли 20 млн человек. В США в зиму 1968-1969 годов 50 млн человек перенесли грипп, 70 тыс. из них скончались.
Наиболее распространенные вирусные инфекции, пути заражения и меры профилактики
либо инфекции, вызываемые как бактериями так и вирусами
Эпидемия — прогрессируемое во времени и пространстве инфекционное заболевание.
Пандемия — инфекционное заболевание захватывающее большие территории (мирового значения). В настоящее время к ряду таких заболеваний относится коронавирусная инфекция (COVID-19) вызванная коронавирусом (SARS-CoV-2).
Профилактика и методы борьбы с вирусами
Основные методы борьбы с вирусными инфекциями — профилактические прививки (вакцины), Ослабленные возбудители болезни, введенные в организм, позволяют выработать иммунитет. Благодаря вакцинам исчезло такое опасное вирусное заболевание, как оспа. Следует помнить, что без оболочки (капсида) вирусная НК сама попасть в клетку не может. Поэтому дезинфекция, вызывающая разрушение белков оболочки вируса (кипячение, хлорирование, обработка карболовой кислотой и др.), — эффективное профилактическое мероприятие. Наш организм тоже обладает защитными механизмами. Так, иммунный белок интерферон способен защищать организм человека от проникновения вирусов гриппа. В целях профилактики воздушно-капельных вирусных инфекций эффективно обрабатывать защитными средствами носовую полость.
Кроме того, сейчас создано несколько видов антивирусных препаратов как на основе неорганических веществ (ремантадин), так и на основе синтетических антител (виферон, биферон и т. д.). Несмотря на то что фармакология и вирусология ведут постоянные успешные исследования, не надо забывать, что соблюдение мер личной гигиены является надежным методом профилактики вирусных инфекций.
Происхождение вирусов
Ученые полагают, что вирусы и бактериофаги представляют собой обособившиеся генетические элементы клеток, подвергшиеся эволюции вместе с клеточными формами жизни.
ВИРУСЫ
Вирусы (лат. virus яд) — неклеточные формы жизни, обладающие собственным геномом и способные к воспроизведению лишь в клетках более высокоорганизованных существ. Для Вирусов в целом характерны две формы существования: внеклеточная, или покоящаяся, и внутриклеточная, размножающаяся (репродуцирующаяся), или вегетативная. Синонимами первого названия являются также термины «вирусная частица», «вирусный корпускул», «вирион», синонимами второго — «комплекс вирус—клетка».
Вирусы существуют в природе, передаваясь от одного хозяина к другому. Репродукция Вирусов может происходить только внутриклеточно, в связи с чем Вирусы являются облигатными внутриклеточными паразитами животных, растений, насекомых, бактерий, грибов и других классов живых существ. Внутриклеточный паразитизм Вирусов обусловлен тем обстоятельством, что они в силу крайней простоты своей организации используют для своего воспроизведения клеточный синтетический аппарат (рибосомы, мембраны), ферменты и энергогенерирующие системы. Связь между двумя формами существования В. осуществляется через нуклеиновую к-ту вириона, к-рая индуцирует в зараженной клетке вирусоспецифические синтезы и, в конечном счете, формирование дочерних вирусных частиц. Различные виды В. на внеклеточной стадии существования характеризуются сравнительно большой вариабельностью размеров (от 15—18 до 3000—3500 нм). Наиболее крупные сложно устроенные В. из рода поксвирусов (Poxvirus), к к-рому принадлежат возбудители оспы и В. осповакцины, различимы в обычном световом микроскопе, но большая часть В. обладает субмикроскопическими размерами и различима лишь в электронном микроскопе.
Содержание
- 1 Химический состав вирионов и свойства вирусных компонентов
- 2 Структура вирусных частиц
- 3 Серологические свойства вирусов
- 4 Комплекс вирус — клетка и основные закономерности репродукции вирусов
- 5 Классификация и номенклатура вирусов
- 6 Морфогенез вирусов
- 7 Генетика вирусов позвоночных
- 7.1 Из дополнительных материалов (том 29)
- 8 Вирусы беспозвоночных
- 9 Вирусы растений
- 10 Таблица. Классификация вирусов по данным 1975 г.
- 11 Классификация вирусов человека и животных (по данным Международного комитета по таксономии вирусов, 1982 г.)
Химический состав вирионов и свойства вирусных компонентов
Вирусные частицы характеризуются простотой своего хим. состава, хотя между различными представителями царства вирусов имеются значительные различия. По этому признаку В. в целом разделяются на две большие группы: простые и сложные. В типичном случае первые В. состоят только из белка и нуклеиновой к-ты, тогда как В. сложные наряду с этими компонентами содержат в своем составе липиды и углеводы, как правило, в форме гликопротеидов.
В свою очередь в пределах этих двух групп также существуют градации. Так, в пределах первой группы В. можно выделить две подгруппы, различающиеся по степени сложности своего белкового компонента. Одна подгруппа образована В., белок которых состоит из одного вида пептидных цепей. К таковым относится большая часть В. растений, и в частности В. табачной мозаики (род Tobamovirus), белок к-рого состоит из 2320 идентичных полипептидов, образованных 158 аминокислотами. Во вторую подгруппу входят В., белок которых образован несколькими типами полипептидных цепей. Различные представители этой подгруппы в свою очередь отличаются по количеству и виду полипептидных цепей. Наиболее просто организованные В. животных, принадлежащие к роду Parvovirus, содержат три вида полипептидных цепей, напр, аденосателлитные В., латентные В. Килхема и др. У сложно устроенных В. типа осповакцины (род Poxvirus) в вирионе насчитывается до 20 различных белков, в т. ч. собственные ферменты. Как правило, сложность состава белка коррелирует со сложностью структурной организации вириона.
Т. о., в типичном случае вирионы представляют собой нуклеопротеиды, т. е. комплекс белка и нуклеиновой к-ты. Исключением из этого правила являются различные дефектные мутанты В., как естественные, так и искусственно полученные, которые вообще не способны синтезировать собственный белок или синтезируют дефектный белок, неспособный образовать нормальную оболочку вирусной частицы. К числу таких дефектных В. можно отнести варианты Вируса погремковости табака, некоторые штаммы В. табачной мозаики, которые в естественных условиях в зараженных клетках существуют в форме комплекса с клеточными белками, а также дефектные варианты В. саркомы Рауса, неспособные синтезировать свою наружную оболочку, формирование к-рой обеспечивается вирусом-помощником (В. птичьего лейкоза).
Вирусы в противоположность клеточным формам жизни содержат в составе вирионов лишь один из двух типов нуклеиновой к-ты: РНК или ДНК, представляющих собой геном вирусной частицы. Т. о., РНК, за к-рой в клетке закрепились лишь функции структурно-метаболические, у В. может выполнять, как и ДНК, функции генетические.
Для В. характерно большое разнообразие форм нуклеиновых кислот, в т. ч. наличие таких форм РНК и ДНК, которые отсутствуют у клеточных форм жизни.
При этом первичная структура РНК и ДНК других В. не обнаруживает каких-либо аномалий по сравнению с клеточными формами нуклеиновых к-т. Однако у многих
В., содержащих ДНК, последняя представлена не линейной, а ковалентно-замкнутой кольцевой молекулой. Такая форма ДНК обнаружена у представителей родов полиомавирусов (Polyomavirus), папиллома-вирусов (Papillomavirus) и липовирусов (Lipovirus).
Из аномалий, связанных со вторичной структурой нуклеиновых кислот В. и не встречающихся среди клеточных нуклеиновых кислот, интересно отметить существование однонитчатых ДНК и двухспиральных РНК. Первая обнаружена у представителей родов Bullavirus и Inovirus, а также среди парвовирусов (Parvovirus): мелкого В. мышей, латентного В. крыс Килхема и аденосателлитных В. Двухспиральная РНК присутствует у реовирусов (род Reovirus) и В. раневых опухолей растений, карликовости риса и цитоплазменного полиэдроза, родовая принадлежность которых пока точно не установлена.
Нуклеиновые кислоты практически всех просто организованных В. обладают инфекционностью. При заражении чувствительных хозяев депротеинизированными препаратами ДНК или РНК этих В. наблюдается типичный инфекционный процесс, заканчивающийся формированием зрелых вирионов потомства. Спектр хозяев и клеток, зараженных препаратами инфекционных нуклеиновых кислот, шире по сравнению с естественным кругом хозяев данного В., т. к. в этом случае нет ограничений, накладываемых специфическим взаимодействием рецепторов В. и клетки. Инфекционность отсутствует у препаратов РНК, выделенных из сложных В. (миксо-, парамиксо-, Рабдовирусы).
Некоторые Вирусы, неспособные синтезировать белки своей оболочки, могут существовать даже в естественных условиях в форме свободной РНК. Обнаруженные этиологические агенты, вызывающие веретеновидность клубней картофеля и экзокортисное заболевание цитрусовых и получившие специальное название «вироиды», представляют собой свободные низкомолекулярные РНК, сходные по размерам с тРНК или 5sPHK. Заражение растений картофеля и цитрусовых препаратами этой РНК вызывает типичную картину заболеваний и репликацию вироидной РНК.
Липидный компонент В. исследован сравнительно хорошо лишь у ортомиксовирусов, парамиксовирусов и арбовирусов (роды Orthomyxovirus, Paramyxovirus, Fiavivirus и Alphavirus). Источником происхождения липидов в этом случае является клеточная мембрана, откуда В. в процессе созревания заимствует свой липидный компонент. Глико-протеиды входят в состав поверхностных образований вирусных частиц, напр. гемагглютининов у миксовирусов и парамиксовирусов. Пути их синтеза пока не изучены.
Структура вирусных частиц
Белок вирионов выполняет двоякую функцию. Во-первых, он образует наружную оболочку (капсид), защищающую нуклеиновую к-ту вирусной частицы. Нуклеиновая к-та с окружающим ее белком обозначается термином «нуклеокапсид», или «нуклеоид». Последний термин обычно применяется морфологами для обозначения внутренних структурных образований, содержащих нуклеиновую к-ту у сложно устроенных В. позвоночных. У просто организованных В. термины «нуклеокапсид» и «вирион» по существу тождественны, но у многих сложно устроенных В. наряду с белковым капсидом имеется еще одна или несколько внешних оболочек, чаще всего липидных или белковых, для обозначения которых пользуются термином «суперкапсид». Так, у В. гриппа имеется внутренний нитевидный нуклеопротеид (нуклеокапсид), известный под названием внутреннего антигена, или S-антигена, заключенный внутри белковой базальной мембраны и внешней липопротеиновой оболочки (суперкапсид).
Для выполнения своих защитных функций капсид В. должен обладать достаточно высокой стабильностью, особенно по отношению к таким факторам внутриклеточной среды, как деструктивные ферменты. В связи с этим нативные неповрежденные вирионы устойчивы к высоким концентрациям протеолитических ферментов. Исключение из этого правила составляют лишь немногие, сложно устроенные В., как, напр., арбовирусы группы Б (род Flavivirus). Эта устойчивость обусловлена высокоупорядоченной третичной и четвертичной структурой белковых субъединиц (капсомеров) в составе капсида, благодаря чему пептидные связи, чувствительные к протеазам, стерически недоступны для действия ферментов. После частичной денатурации или деградации белковый капсид В. легко гидролизуется протеазами.
Вторая функция капсида заключается в обеспечении адсорбции В. только на тех клетках, где может происходить их размножение. Такая специфичность взаимодействия В. с ограниченным кругом хозяев обеспечивается наличием на поверхности вирионов специфических рецепторов, которые соответствуют определенным рецепторам на поверхности клеток. В тех случаях, когда В. лишены специализированных рецепторов, как, напр., В. растений, их попадание в клетки природных хозяев обусловливается определенными переносчиками.
К числу простых относятся все Вирусы, представляющие собой «голый» нуклеокапсид, хотя их оболочка может состоять из нескольких видов полипептидных цепей. В этом случае структура вирусных частиц подчиняется строгим правилам симметрии (см. Симметрия). У сложно устроенных В., имеющих высоко-дифференцированную структуру и суперкапсиды, симметрия, как правило, становится более сложной.
Серологические свойства вирусов
Поскольку Вирусы содержат белки, а в ряде случаев гликопротеиды и липопротеиды, то вирионы несут определенные антигенные детерминанты, вызывающие образование специфических антител как при естественных формах инфекции, так и в опытах по иммунизации лабораторных животных. Просто устроенные В., содержащие один вид белка, вызывают образование одного вида антител, сложно устроенные В., содержащие различные виды белков и разные структурно-морфологические компоненты, несут несколько типов антигенов.
Внеклеточные В. при контакте с антителами против наружных антигенов теряют свою инфекционность (нейтрализуются), но из неинфекционного комплекса вирус—антитело снова можно получить инфекционные вирионы после диссоциации такого комплекса при воздействии кислых pH, температуры и высокой ионной силы. В. внутриклеточные к нейтрализующему действию анти-сыворотки устойчивы, и такой прием нередко используется для изучения адсорбции и проникновения В. в клетки.
По вопросу о роли антител в иммунитете к вирусным инфекциям единой точки зрения нет, что, по-видимому, связано с различиями в биологии разных В., путей их распространения и циркуляции в организме, местах синтеза, доступности для антител и т. п. (см. Иммунитет противовирусный).
Комплекс вирус — клетка и основные закономерности репродукции вирусов
Для В. в целом характерен так наз. дизъюнктивный (лат. disjunctus разобщенный) путь репродукции, означающий, что в процессе репликации В. родительский вирион как таковой исчезает.
В зараженной клетке присутствие вирусных частиц не обнаруживается ни с помощью серологических, ни с помощью электронномикроскопических методов вплоть до поздних стадий инфекции, когда появляются первые дочерние вирионы (так наз. латентный период инфекции, синонимами к-рого являются термины «эклипс-период», «период маскировки», фаза смены информаций — «си-фаза»).

Проникновение В. в клетку происходит по механизму, близкому к пиноцитозу (см.), но конечный результат остается тем же (рис. 1). В этом случае инфицирующий вирион после адсорбции на клеточных рецепторах захватывается клеточной мембраной, к-рая впячивается внутрь клетки, образуя вакуоль. В процессе взаимодействия вириона с клеточной мембраной его капсид претерпевает деструктивные изменения и приобретает чувствительность к клеточным протеазам, которые и разрушают вирусный белок, освобождая вирусную нуклеиновую к-ту. У сложно устроенных В. из родов Poxvirus, Paramyxovirus, Orthomyxovirus и Rhabdovirus течение этого процесса осложняется тем, что удаление суперкапсида и капсида нуклеоида разобщено во времени и проходит в две стадии. При этом нуклеиновая к-та может быть транскрибирована непосредственно в составе нуклеокапсида или нуклеоида, освободившихся от суперкапсида. Процесс транскрипции в этом случае осуществляется с помощью вирусных РНК-полимераз, входящих в состав нуклеоида (см. Транскрипция).
Следующая особенность дизъюнктивного способа репродукции В. состоит в том, что синтез элементов вирусного корпускула осуществляется в виде пулов (обменных фондов), которые разобщены и в пространстве, и во времени. Нуклеиновая к-та вируса может реплицироваться, напр., в ядре или ядрышке, белок может синтезироваться в цитоплазме, а сборка целых вирионов или нуклеокапсидов может происходить на внутренней поверхности цитоплазматической мембраны. Наконец, сложный липопротеиновый суперкапсид может приобретаться В. в процессе отпочкования.
Последующей стадией репродукции (после проникновения вирусной нуклеиновой к-ты в клетку) является синтез вирусоспецифических ферментов и белков, необходимых для репликации В. (так наз. ранние белки, не входящие в состав вирионов). Детали этого процесса у разных В. различаются в зависимости от того, какой тип нуклеиновой к-ты входит в состав вирионов. У большей части В., которые содержат однонитчатую РНК, последняя способна выполнять одновременно и функции генетической детерминанты, и функции собственной информационной РНК — так наз. (+) цепи. Эти родительские (+) цепи РНК непосредственно связываются с рибосомами и транслируются, т. е. наводят в них синтез вирусоспецифических белков. В этом случае первым актом вирусоспецифических синтезов в зараженной клетке будет трансляция вирусной (+) матрицы. У В., содержащих ДНК как однонитчатую, так и двунитчатую, а также однонитчатую РНК, неспособную к трансляции (—) цепи, или двунитчатую РНК, первым синтетическим актом должно быть формирование комплементарной полинуклеотидной цепи, т. е. синтез вирусоспецифических ранних информационных РНК.
Естественно, что этот процесс должен осуществляться с помощью вирусных либо клеточных РНК-полимераз. Если в клетке имеются ДНК-зависимые РНК-полимеразы, способные транскрибировать вирусную ДНК, то РНК-зависимых РНК-полимераз в нормальной клетке нет. В связи с этим последний фермент для осуществления первичного акта транскрипции РНК должен проникать в клетку вместе с инфицирующей вирусной РНК, как правило, в составе нуклеоида или нуклеокапсида. В дальнейшем, после трансляции инфекционных РНК в зараженной клетке появляются и дочерние, т. е. вновь синтезированные РНК-зависимые РНК-полимеразы.

На третьей стадии вирусной репродукции происходит репликация вирусной нуклеиновой к-ты, к-рая в случае двухспиральных нуклеиновых кислот осуществляется по известному симметричному полуконсервативному механизму, а в случае однонитчатых нуклеиновых кислот — по асимметричному полуконсервативному механизму, через стадию репликативной (РФ) и репликативной промежуточной (РПФ) форм. Как видно из схемы, представленной на рис. 2, на матрице родительской (+) цепи синтезируется комплементарная ей (—) цепь, в результате чего возникает двухспиральная РФ.
Затем на матрице (—) или (+) цепей начинается одновременный синтез нескольких (+) или (—) полинуклеотидных цепей, соответственно и возникает РПФ.
Четвертая стадия репродукции — это синтез информационных РНК, кодирующих поздние белки, к-рыми являются конститутивные белки, входящие в состав вирионов. Одновременно начинается и формирование вирионов.
Как показали работы Шрамма (G. Schramm) и Фреккель-Конрата (H. Fraenkel-Conrat), индивидуальные полипептидные цепи таких простых В., как В. табачной мозаики, при нейтральных или слабокислых значениях pH способны спонтанно агрегировать в растворе, образуя вирусоподобные частицы (белковые капсиды). При наличии в растворе нуклеиновой к-ты этих В. происходит автоматическая сборка инфекционных вирусных частиц, протекающая как чисто физ.-хим. реакция агрегации и не требующая участия дополнительных факторов. Аналогичным образом происходит и сборка вирионов в зараженных клетках после того, как концентрация вирусных РНК и белка в соответствующих пулах достигнет критического уровня. У сложно устроенных В. принципы самосборки сохраняются лишь на уровне отдельных компонентов вирусных частиц, тогда как сборка этих компонентов в единое целое требует участия многих дополнительных факторов и ферментов, которые сами по себе в состав зрелых вирионов не включаются. Такого рода морфопоэтические факторы В. синтезируются на этой стадии инфекции вместе с конституционными вирусными белками. (См. ниже Морфогенез вирусов.)
Заключительная стадия вирусной инфекции, смыкающаяся с формированием зрелых вирионов, заканчивается освобождением дочерних вирусных частиц в среду. В зависимости от структурной организации и сложности вирионов, а также биологии данного В. и его взаимоотношения с клеткой хозяина этот процесс может осуществляться по-разному у разных групп В. Вирусы могут либо тем или иным способом лизировать клетки, обусловливая цитопатическое действие (патогенные, или литические, вирусы), либо освобождаться из клетки, не разрушая ее (бессимптомная инфекция). У литических, патогенных В. позвоночных и беспозвоночных разрушение клетки достигается либо за счет активации лизосом, либо путем интенсивного повреждения клеточных мембран в процессе отпочкования.
Многие вирусные штаммы и даже виды обладают тем или иным генетическим дефектом и способны репродуцироваться лишь в присутствии других В.— помощников. Классическими примерами являются: штамм Брайена В. саркомы Рауса, реплицирующийся лишь в присутствии В. птичьего лейкоза; аденосателлитный В., способный размножаться в присутствии полноценного аденовируса, мелкий сателлитный Вирусов некроза табака, нуждающийся для успешной репродукции в присутствии крупных частиц В. некроза табака. ДНК аденосателлитного В. и РНК В. некроза табака содержат информацию для синтеза собственного капсидного белка и не содержат полной информации для репликации в клетке.
Наряду с этим имеется также и весьма большая группа РНК-содержащих В. растений с фрагментированным геномом, который распределен среди нескольких видов частиц, образующих инфекционный препарат лишь при наличии всех компонентов.
Так, инфекционный препарат В. мозаики коровьего гороха состоит из трех типов вирионов с коэффициентами седиментации 58S, 95S и 115S, В. мозаики люцерны — из пяти различных компонентов с коэффициентами седиментации 99S, 83S, 76S, 68S, 61S и 53S. Многокомпонентными системами являются также В. мозаики костра, В. стрика табака и некоторые другие. Каждый из компонентов этих В. с фрагментированным геномом сам по себе не инфекционен и содержит лишь определенную часть генетической информации, необходимой для успешного функционирования всей системы в целом.
Взаимоотношения между вирусами и клетками отличаются большой сложностью и определяются как типом вируса, так и геномом клетки и ее физиологическим состоянием.
Вирусный геном в зараженной клетке может вступать в определенные генетические отношения с геномом клетки. По этому признаку В. можно разделить на две большие группы: с автономной репликацией геномов (инфекционные вирусы) и В., способные тем или иным образом взаимодействовать с клеточным геномом (интеграционные вирусы). Последняя группа включает опухолеродные В., нуклеиновая к-та которых способна встраиваться (интегрироваться) тем или иным образом в клеточную хромосому в форме провируса, вызывая трансформацию клеток (см. Трансформация). Границы между этими группами весьма условны, и один и тот же В. в зависимости от вида клеток может вести себя либо как инфекционный, либо как интеграционный. В. с автономной репликацией геномов в свою очередь могут быть по отношению к одним хозяевам патогенными, а по отношению к другим хозяевам нейтральными. Все интеграционные В. в той или иной мере вступают в симбиотические отношения с клеткой хозяина.
Литический, или патогенный, путь развития инфекции определяется тем, что клетки, где происходит репродукция В., в конечном счете погибают.
Летальный исход инфекции для клетки может вызываться рядом независимых причин: 1) специфической и необратимой блокировкой деятельности клеточного генома на ранних стадиях инфекции; 2) неспецифическими повреждениями клеточного генома в процессе инфекции; 3) переключением метаболических ресурсов клетки на вирусоспецифические синтезы; 4) специфическим лизисом клетки, связанным с необходимостью освобождения дочерних вирусных частиц в среду; 5) нарушением структуры клеточных мембран в результате интенсивного выхода вирусных частиц в среду, особенно в процессе отпочкования, и некоторыми другими.
Как видно из этого перечня, лишь первая и третья причины связаны со специфическим цитотоксическим или цитопатогенным действием, запрограммированным в вирусном геноме и реализующимся в процессе инфекции. Остальные из перечисленных причин, приводящих к гибели клеток, строго не детерминированы и обусловлены в основном интенсивностью вирусоспецифических синтезов. Поэтому при некоторых ограничениях в скорости синтеза и освобождения зрелых вирусных частиц в среду клетка может сохранять свою жизнеспособность и длительное время продуцировать вирусные частицы. Несомненно, что такой нейтральный тип взаимоотношений, лежащий в основе бессимптомных инфекций, безусловно, выгоден для В. как вида и представляет собой наименьшее зло для хозяина.
Наконец, между В. и клетками могут существовать и вполне симбиотические отношения, когда несомненные выгоды получают оба партнера комплекса. Примеры такого рода можно найти у онкогенных В. Как известно, трансформация клеток онкогенными В. создает также помехи для суперинфекции в виде феномена интерференции (см. Интерференция вирусов).
Выгоды, которые получают В. при таком интеграционном пути развития инфекции, также несомненны. Прежде всего репликация нуклеиновой к-ты В. целиком обеспечивается клетками, поскольку провирус является частью клеточной хромосомы. Поэтому каждая дочерняя клетка, образовавшаяся в результате деления родительской клетки, несет в себе провирус, что обеспечивает вертикальную передачу В. и его распространение в пределах данного вида хозяев.
Классификация и номенклатура вирусов
В 1966 г. на IX Международном конгрессе микробиологов в Москве был создан Международный комитет по номенклатуре вирусов (МКНВ), который был позже переименован в Международный комитет по таксономии вирусов (МКТВ). МКТВ является органом вирусол. секции Международной ассоциации микробиологов и состоит из представителей национальных обществ. При МКТВ образованы подкомитеты по
В. позвоночных, беспозвоночных, растений и бактерий, которые имеют в своем составе так наз. группы изучения отдельных разделов; напр., в подкомитете В. позвоночных имеются группы изучения энтеровирусов, реовирусов, арбовирусов и др.
Сбор информации о В. и систематизация ее осуществляются в соответствии с характеристикой В. по следующим основным показателям: а) нуклеиновой к-те, белкам, липидам, углеводам, морфологии и физ.-хим. свойствам; б) репликации; в) феноменам генетических взаимодействий; г) кругу хозяев; д) патогенности; е) географическому распространению; ж) способу передачи; з) антигенным свойствам. Значимость каждого из показателей в построении иерархической системы пока еще не до конца определена, но, по-видимому, первая послужит основой формирования крупных таксономических групп (семейства и выше).
Общность антигенной структуры является обязательным признаком при определении вида.
В современной классификации В. (таблица) рассматриваются как единая система, независимо от круга основных хозяев (позвоночных, беспозвоночных, растений, бактерий). Предполагается создание полной иерархической системы, подобной классификации животных, включающей такие таксономические группы, как вид, род, семейство, порядок, класс, и, возможно, еще более высокие категории. Пока не удалось достигнуть четкого определения понятий «вид» и «род». Под видом разумеют группу В. с идентичными свойствами, а род представляет собой группу В., сходных по ряду общих свойств.
К В. применяется биноминальная номенклатура, в соответствии с к-рой наименование вида состоит из родового и видового названий. В силу сложившихся привычек в номенклатуре В. в основном сохраняются уже существующие названия, в т. ч. буквенные и цифровые, если они общеупотребительны. Новые названия создаются по правилам словообразования в латинском и греческом языках. Название порядков оканчивается на «-ales», семейств — на «-idae», родовое название включает слово «virus». Название семейств и родов пишется с прописной буквы, видовое — со строчной. Для некоторых определяющих свойств В. применяется кодированная запись в виде криптограммы (надпись, сделанная знаками), что облегчает восприятие свойств каждого В. или отдельных групп и сопоставление их между собой.
Криптограмма состоит из четырех пар символов, разделенных двоеточием, имеющих следующее значение:
1. Тип нуклеиновой к-ты / число нитей.
2. Молекулярный вес нуклеиновой к-ты / процентное содержание в вирионе.
3. Внешние очертания вириона / очертания нуклеокапсида.
4. Хозяин / переносчик.
Для выражения свойств первой пары употребляют следующие символы: РНК — R, ДНК — D, однонитчатая — 1, двунитчатая — 2.
Молекулярный вес выражают в миллионах дальтон. Если нуклеиновая к-та фрагментирована и состоит из нескольких кусков, то в случае, когда различные куски находятся вместе в одном типе частиц, указывают общий мол. вес со знаком 2. Если же фрагменты нуклеиновой к-ты находятся в различных частицах, то состав и мол. вес в каждой перечисляются отдельно.
Форму вириона и нуклеокапсида обозначают: S — сферическая; E — продолговатая с параллельными сторонами, концы не закруглены; U — продолговатая с параллельными сторонами и закругленным(и) концом(ами); X — комплексная структура.
Для обозначения хозяина используют: А — актиномицеты; В — бактерии; F — грибы; I — беспозвоночные; P — птеридофиты (Pteridophyte), споровые растения; S — семенные растения; V — позвоночные; для обозначения переносчика: Ас — клещики и клещи (Acarina, Arachnida); Al — белокрылки (Aleyrodidae, Hemiptera, Insecta); Ap — тли (Aphididae, Hemiptera, Insecta); Au — цикадовые (Auchenorrhyncha, Hemiptera); Cc — мучной червец (Coccidae, Hemiptera); Cl — жуки (Colleoptera, Insecta); Di — мухи и комары (Diptera, Insecta); Fu — грибы (Chytridiales и Plasmodiophorales, Fungi); Gy — мириды, пиезмы, кружевницы (Gymnocerata, Hemiptera); Ne —нематоды (Nematoda); Ps — листоблошки (Psillidae, Hemiptera); Si — блохи (Siphonaptera, Insecta); Th — трипсы (Thysanop-tera, Insecta); Ve — переносчик неизвестен; О — распространяется без переносчиков; * — свойства неизвестны. В скобки ставят данные сомнительные или неподтвержденные.
Для примера приводим криптограмму рода Rhabdovirus, R/1 : 4/2 : U/U:V,I, S/O, Ac, Ap, Di, к-рая расшифровывается так: однонитчатый РНК-содержащий вирус, мол. вес РНК—4 млн. дальтон, что составляет 2% веса вириона. Наружные очертания В. и нуклеокапсида — продолговатые с закругленным концом. Для разных представителей хозяевами являются позвоночные, беспозвоночные, семенные растения. Распространяются через окружающую среду, а также с помощью переносчиков клещей, тлей, двукрылых.
Формирование отдельных таксономических групп идет в зависимости от полноты информации и пока находится на стадии образования родов и семейств. Предложения об образовании таксономических групп и их номенклатуры, исходящие от официально созданных групп изучения или отдельных ученых, рассматриваются соответствующими подкомитетами и приобретают законную силу лишь после утверждения МКТВ.
Морфогенез вирусов
В. проходят сложный строго специфичный для каждой группы В. онтогенетический цикл развития. Морфогенез, или морфопоэз, В. представляет собой основной и наиболее важный этап его онтогенетического цикла развития, который состоит из комплекса последовательных формообразовательных (морфогенетических) процессов, приводящих к образованию вириона — заключительной формы развития В. Процесс индивидуального развития и репродукции В. контролируется его автономной генетической системой — макромолекулярной ДНК или РНК. Геном простейших В. состоит из 1—2 генов, геном сложных ДНК-содержащих В.— из 100 и более генов. Сборка вирионов — заключительный этап морфогенеза В., представляет собой процесс упорядоченной агрегации структурных вирусных компонентов (макромолекул нуклеиновых кислот, структурных белков и т. д.). Сборка вирионов сложных В. регулируется так наз. морфогенетическими генами. Вирионы простейших В. (напр., В. табачной мозаики) или отдельные компоненты сложных вирионов формируются в результате самосборки, к-рая представляет собой процесс спонтанной упорядоченной молекулярной агрегации и определяется в основном структурой белковых субъединиц. Существует определенная зависимость между степенью сложности архитектуры вирионов и сложностью их морфогенеза, т. е. чем сложнее организация вириона, тем больший путь дифференцировки проходит В. в онтогенетическом цикле развития и тем большее число морфогенетических генов контролирует этот процесс.



Структура вириона является специфическим признаком каждой группы В. (рис. 3). Вирионам ДНК- и РНК-содержащих вирусов или отдельным их компонентам могут быть присущи два основных типа симметрии: спиральная (винтовая — рис. 4,2—4) и кубическая (рис. 4,2 и 5 и рис. 5 1—3). У некоторых сложно организованных вирионов обнаружен комбинированный тип симметрии (рис. 4, 6 и 7 и рис. 5, 4 и б).
Спиральная укладка макромолекул характеризуется шагом спирали, величиной винтового смещения, длиной спиральной цилиндрической частицы, а также числом субъединиц на один оборот спирали. Если форма белковых субъединиц приближается к сфере, то при их спиральной укладке вдоль оси спирали образуется полость (канал). Подобную организацию имеют все нуклеокапсиды и рибонуклеопротеидные тяжи со спиральной симметрией.
Вирионы с кубическим типом симметрии обнаружены у многих В. человека, животных, растений и бактерий. Эти вирионы характеризуются величиной, группой симметрии, числом и типом осей вращения, числом и морфологией капсомеров. Большая часть вирионов с кубическим типом симметрии построена по типу двадцатигранника (икосаэдра), который имеет оси симметрии второго, третьего и пятого порядков. Кубический тип симметрии характерен или для всего вириона, или для внутреннего компонента сложных вирионов.
В 1967—1970 гг. было показано (А. А. Авакян и А. Ф. Быковский), что цикл развития каждого В. состоит из пяти стадий, к-рым соответствуют пять форм существования В.
Т. о., цикл развития В. можно представить следующим образом. Проникший в чувствительную клетку виронуклеон индуцирует образование полигеномного виропласта, который представляет собой комплекс структур, обеспечивающих репликацию нуклеиновой к-ты В., синтез специфических антигенов, а также скопления этих компонентов. Виропласты обнаружены в цикле развития всех ДНК- и многих РНК-содержащих В. человека и животных.
Некоторые из них локализуются в цитоплазме (В. группы оспы, реовирусы, энтеровирусы), другие — в ядре инфицированной клетки (В. группы герпеса, паповавирусы и др.). Конечный этап дифференцировки полигеномного виропласта — формирование моногеномных виропластов, каждый из которых является начальной стадией онтогенеза нового поколения В. и включает в себя геном В., набор специфических ферментов и структурных белков. Развитие моногеномного виропласта завершается синтезом (формированием) de novo оболочки В. Затем В. (провирион) отделяется от виропласта, и последующая дифференцировка его структуры идет в морфологически изолированной от внешней среды (органоидов клетки) полости. При этом наблюдается строгая последовательность и преемственность морфогенетических процессов.
В результате сборки и самосборки происходит не только увеличение массы структурных компонентов провириона до определенной (строго постоянной для каждого вида В.) величины, но и наблюдается последовательное чередование роста и дифференцировки структур В. По-видимому, окончание формирования ряда структур (напр., первичной оболочки В. оспы) является своеобразным сигналом, включающим следующий этап дифференцировки провириона. В результате этих динамических морфогенетических процессов образуется вирион — споровая форма В., обеспечивающая сохранение генома, следовательно, и вида В. После освобождения генома В. из вириона начинается новый цикл развития В.
Морфогенез простейших РНК-содержащих Вирусов можно описать как процесс самосборки. Это характерно не только для такого простого В. как спутник В. табачного некроза, геном к-рого — молекула РНК — имеет мол. вес ок. 400 000 дальтон и может кодировать только один белок, но и для В. табачной мозаики, геном к-рого состоит из 5—6 генов. Более сложен морфогенез у РНК-содержащих В. животных и человека, вирионы которых, кроме нуклеокапсида, имеют внешнюю оболочку (миксовирусы, онкорнавирусы, арбовирусы и пр.).



Геном таких В. является полицистронным и программирует синтез не одного-двух, а многих белков, в т. ч. и тех, которые не входят в структуру вириона. Синтез РНК некоторых из этих В. происходит в цитоплазме, других — в ядре клетки; синтез же вирусных белков идет только в цитоплазме на полирибосомах в зоне виропласта. Отдельные компоненты этих В., напр, рибонуклеопротеидные тяжи миксовирусов, формируются в процессе самосборки, однако композиция вириона требует более сложных морфогенетических процессов, в субмикроскопической морфологии которых выявляется ряд последовательных этапов: проникновение в клетку, внутриклеточный морфогенез и выделение из клетки (рис. 6—8).
В процессе морфогенеза тогавирусов, миксовирусов, онкорнавирусов, рабдовирусов в состав вирионов входят компоненты мембранных клеточных структур (цитоплазматической оболочки, эндоплазматического ретикулума, ядерной оболочки, комплекса Гольджи, мембран митохондрий), модифицированных в процессе инфекции.


Морфогенез ДНК-содержащих В. также весьма различен у разных групп этих В. и зависит от сложности архитектуры вириона (рис. 9 и 10).
Морфогенез В., так же как и структура вириона, является специфическим признаком каждой группы В.
Генетика вирусов позвоночных
Геном В. позвоночных может быть представлен различными формами нуклеиновых кислот (см. выше).
В структуре генома В. позвоночных, как и других живых организмов, различают отдельные участки: гены, цистроны, мутоны, реконы. В геномах В. позвоночных может быть закодировано от 6—8 (пикорнавирусы) до нескольких сотен (поксвирусы) белков. Попытки построить генетические карты В. позвоночных пока не дали положительных результатов. Лишь в опытах с полиовирусом удалось установить, что на 51 конце РНК располагается генетическая информация на капсидные белки вириона, и определить последовательность цистронов, кодирующих эти белки [Рекош (D. Rekosh)].
Свойства В., передающиеся по наследству, называются генетическими признаками. Совокупность всей наследственной информации В. определяет его генотип (см.). В результате взаимодействия генотипа и окружающей среды формируется фенотип, под к-рым понимается совокупность всех проявляемых в конкретных условиях генетических признаков. Среди генетических признаков В. позвоночных чаще всего изучают вирулентность, характер и размер бляшек, образуемых В. на культуре ткани под агаром, характер и размер поражений на хорионаллантоисной мембране куриных эмбрионов, способность размножаться при измененной температуре. Изучают также такие генетические признаки, как характер цитопатических изменений, способность к размножению в различных видах клеток, способность индуцировать образование интерферона и чувствительность к нему, способность образовывать бляшки при особых условиях (измененная концентрация соды, в присутствии декстрана и пр.), размножение в присутствии ряда ингибиторов и пр. Изучают также генетические признаки, отражающие особенности вириона, напр, морфологию вириона, константу седиментации его, устойчивость инфекционности или других биол, свойств вириона к температуре, ультрафиолетовым лучам и т. п., способность к адсорбции на биол, и хим. субстанциях (эритроциты, ДЭАЭ-целлюлоза и пр.) и т. д. Предполагается, что отдельный генетический признак В. связан с функцией какого-либо одного вирусоспецифического белка. Необходимо учитывать, однако, что мутационные повреждения в одном участке генома нередко приводят к изменению нескольких генетических признаков, имеющих различное фенотипическое проявление (плейотропия). В то же время в основе изменения генетического признака, имеющего одно и то же фенотипическое выражение (напр., способность размножаться при повышенной температуре), могут лежать мутационные повреждения различных генов.
В., как и другие организмы, способны изменять свои свойства как в естественных условиях размножения, так и в эксперименте. В основе наследственного изменения свойств В. могут лежать два процесса: 1) мутация (см.), т. е. изменение последовательности или состава нуклеотидов в определенном участке генома В., ведущее к фенотипически проявляемому изменению свойства; 2) рекомбинация (см.), т. е. обмен генетическим материалом между двумя близкими, но отличающимися по наследственным свойствам В. Поскольку вирусологи исследуют свойства не отдельных вирусных частиц, а вирусной популяции в целом, для проявления изменения наследственности В. необходим второй процесс — селекция, т. е. возникновение условий, при которых происходит преимущественное размножение вирусных частиц с измененной наследственностью, в результате чего вся вирусная популяция будет состоять из таких вирионов. Частота мутационных повреждений отдельных генов В. в естественных условиях может значительно варьировать. Так, частота изменения u-признака (характер поражений на хорион-аллантоисной мембране куриных эмбрионов) В. оспы коров может достигать 1 : 10 2 — 1 : 10 3 , в то время как мутации по ts-признаку (температурочувствительность) возникают весьма редко (1:10 7 ). Хорошо известным примером изменения наследственных свойств в процессе размножения в естественных условиях является В. гриппа. Следует, однако, отметить, что изменчивость этого В. в последнее время связывают также с возможностью образования в естественных условиях рекомбинантов В. гриппа человека и животных [Уэбстер, Лей-вер (R. Webster, W. Laver, 1973)]. Изменения наследственных свойств В. в эксперименте осуществляются путем индукции мутаций при обработке В. мутагенами, путем рекомбинации, а также с помощью специальных пассажей. При индукции мутаций с помощью мутагенов используют как непосредственную обработку мутагеном нативного В. или вирусной нуклеиновой к-ты in vitro, так и добавление мутагена в систему, в к-рой размножается В. Частота возникновения мутаций при обработке В. мутагенами широко варьирует. Изменения наследственных свойств В. путем специальных пассажей (длительные пассажи при обычных условиях культивирования, пассажи при измененной температуре, в присутствии ингибиторов и пр.) как бы имитируют процесс изменчивости В. в естественных условиях, приводя к селекции мутантов, наиболее приспособленных к размножению в используемых условиях пассажей. Известны случаи, когда размножение В. в определенной чувствительной системе приводит к изменению некоторых свойств В., однако при последующей репродукции В. в иной системе эти изменения очень быстро исчезают. Такого рода изменчивость (модификационная) не является наследственной и обусловлена включением в вирион некоторых компонентов клетки-хозяина.
Получено большое количество мутантов вирусов позвоночных, т. е. вирусов, у которых в результате мутационного повреждения участка генома изменилось определенное наследственное свойство. В последние годы особое внимание привлекают условно летальные мутанты, использование которых позволило изучить ряд важных особенностей строения генома и выявить некоторые новые стадии репродукции. Условно летальные мутанты — мутанты, у которых жизненно важный белок в результате мутационного повреждения утрачивает способность функционировать при определенных неразрешающих условиях, но нормально осуществляет свою функцию в обычных условиях репродукции. У В. позвоночных известны два вида условно летальных мутантов: 1) температурочувствительные (ts) мутанты, неспособные, в отличие от ts+ штаммов (диких), в условиях повышенной температуры образовывать инфекционные вирусные частицы, и 2) зависимые от хозяина hd-мутанты, утерявшие способность размножаться на некоторых видах клеток. Температурочувствительные мутанты получены в опытах с пикорнавирусами, тогавирусами, миксовирусами, рабдовирусами, ретровирусами, реовирусами, паповавирусами, аденовирусами и В. группы оспы; hd-мутанты обнаружены у пикорнавирусов, В. оспы и вируса герпеса.
В процессе одновременного размножения нескольких вирусных частиц в одной клетке между этими частицами могут происходить генетические взаимодействия и обмен генетическим материалом. Известно несколько видов генетических взаимодействий В. позвоночных. Множественная реактивация — процесс, который имеет место при заражении клеток В., частично инактивированными ультрафиолетовыми лучами, при множественности инфекции более одной вирусной частицы на клетку. Поскольку при ультрафиолетовом облучении происходит разрушение лишь отдельных участков генома, в основе феномена множественной реактивации лежит рекомбинация сохранивших нативность участков генетического материала нескольких проникших в клетку вирусных частиц с образованием полноценного генома. Такой феномен удалось наблюдать в опытах с В. гриппа и В. группы оспы. Рекомбинация — обмен генетическим материалом между размножающимися в одной клетке вирусными частицами, различающимися по нек-рым наследственным признакам. В результате образуются рекомбинанты (гибриды), обладающие частью свойств одного, а частью свойств другого родительского штамма (см. Рекомбинация). Рекомбинанты В. позвоночных удается получать только при скрещивании близких по свойствам В., принадлежащих к одной группе. Частота возникновения рекомбинантов широко варьирует и существенно зависит от используемой системы (клетки, вирус), а также от того, какое наследственное свойство стремятся рекомбинировать. Рекомбинация с высокой частотой наблюдается у РНК-содержащих В. (ортомиксовирусов, реовирусов, онкорнавирусов) и у всех ДНК-содержащих В., геном которых представлен двунитчатой ДНК. Кросс-peактивация (спасение маркера) — феномен, сходный с рекомбинацией, однако один из участвующих В. используют в нативном виде, а другой инактивируют путем частичного разрушения генетического материала (ультрафиолетовое облучение, слабое нагревание). С помощью кросс-реактивации удалось получить рекомбинанты при скрещивании В. гриппа и В. оспы. Гетерозиготность — феномен, заключающийся в том, что при одновременном размножении в клетке нескольких частиц В., различающихся по наследственным свойствам, могут образовываться вирионы, содержащие полный геном одного родительского штамма и, кроме того, часть генома (или полный геном) другого В. (диплоидные или полиплоидные вирионы). Хотя такого рода объединение генетического материала в одной вирусной частице не наследуется, оно позволяет такому вириону дать потомство, в к-ром будет содержаться часть вирусных частиц со свойствами одного, а часть — другого родителя. Вирусные частицы, дающие описанный феномен, получили название гетерозигот, в отличие от обычных гомозиготных частиц, все потомство которых обладает одинаковыми свойствами. Феномен гетерозиготности обнаружен в опытах с В. гриппа и В. ньюкаслской болезни. Транскапсидация — феномен, при к-ром часть чужеродного генетического материала, заключенного внутри капсида другого неродственного В., способна переноситься в стабильной форме в чувствительные к основному В. клетки. Наблюдается при одновременном выращивании в клетках аденовируса и обезьяньего вируса SV40, в процессе к-рого часть генетического материала вируса SV40 ковалентно связывается с ДНК аденовируса и при последующем проникновении такой частицы в клетки этот участок генома вируса SV40 способен индуцировать образование трансформирующего антигена.
Из дополнительных материалов (том 29)
Генетика вирусов позвоночных является одним из важнейших разделов вирусологии, в задачу к-рого входит изучение наследственности и изменчивости вирусов человека и животных, а также установление функции вирусных генов и кодируемых ими белков.
Исследования в области генетики вирусов позвоночных значительно интенсифицировались в последние годы. Полученные результаты были использованы не только для понимания механизмов репродукции, строения генома и функции отдельных генов, но и в практической медицине. Так, выявление функции вирусных генов помогло выяснить механизмы патогенности вирусов, их способность преодолевать иммунные барьеры организма. Изучение мутационной и рекомбинационной изменчивости вирусов способствовало выяснению механизмов возникновения новых вариантов вирусов, обладающих резко выраженной вирулентностью, а также резистентностью к противовирусным ингибиторам. Генетические исследования вирусов помогают также установить причины возникновения персистентных и вялотекущих вирусных инфекций, выяснить влияние вируса на геном клетки-хозяина (с одной стороны, интеграция генома вируса в геном клетки как возможная причина возникновения злокачественных опухолей, с другой — повреждения хромосом, ведущие к генетическим заболеваниям). Создание современных живых и инактивированных вирусных вакцин оказалось возможным лишь на основе знания генетики вирусов и освоения методов генетических исследований.
Примерно у 80% известных вирусов генетическим материалом является РНК, а у 20% — ДНК. У РНК-содержащих вирусов геном может быть представлен в нескольких формах: линейной одноцепочечной молекулой РНК, обладающей инфекционными свойствами и способной служить матрицей для синтеза вирусных белков; такой же молекулой РНК, но не обладающей инфекционными свойствами и матричной активностью (так наз. вирусы с негативным геномом); несколькими молекулами одноцепочечной линейной РНК, содержащими сходную генетическую информацию; сегментированной линейной одноцепочечной РНК, большинство сегментов к-рой кодирует один вирусный белок; сегментированной линейной двуцепочечной РНК, каждый сегмент к-рой кодирует один вирусный белок. У ДНК-содержащих вирусов геном может быть представлен одноцепочечной линейной ДНК, двуцепочечной циркулярной ДНК, двуцепочечной линейной ДНК. Обнаружено, что у нек-рых вирусов, геном к-рых представлен как ДНК (паповавирусы), так и РНК (ортомиксовирусы), один и тот же ген может кодировать не один, а несколько белков (в результате сдвига рамки кода с одного гена).
Благодаря развитию генетической инженерии удалось синтезировать ДНК-копии отдельных генов или всего генома РНК-содержащих вирусов позвоночных, вставить (с помощью плазмид) участки вирусных генов в геном кишечной палочки и получить клоны этих бактерий, в к-рых происходит интенсивный синтез ДНК-копий генов РНК-co держащих вирусов или отдельных генов ДНК-содержащих вирусов. Это позволило провести анализ полной последовательности нуклеотидов многих генов и даже всего генома вирусов позвоночных. Применение рестрикционных эндонуклеаз, способных расщеплять нуклеиновые к-ты в строго определенных участках, способствовало построению генетических карт большинства ДНК-содержащих вирусов позвоночных и определению количества генов, их взаиморасположения в геноме, а также выявлению белков, кодируемых конкретным геном, у многих РНК-co держащих вирусов (пикорнавирусы, рабдовирусы, парамиксовирусы, ретровирусы, буньявирусы, ортомиксовирусы, аренавирусы, реовирусы).
Из мутантов вирусов позвоночных, как правило, исследуют условно-летальные мутанты — температурно-чувствительные (ts) и зависимые от хозяина (hd), а также жизнеспособные делеционные мутанты, т. е. мутанты, у к-рых в результате мутации произошло выпадение определенного участка генома. У словно-летальные мутанты вирусов позвоночных получают различными способами: селекцией так наз. спонтанных мутантов; селекцией мутантов из вирусной популяции, обработанной мутагенами; в результате пассажей вирусов в условиях пониженной температуры или в нечувствительной к данному вирусу системе к леток-хозяев.
Значительный прогресс достигнут в построении генетических карт генома ряда вирусов позвоночных. Сегменты генома различных штаммов одного и того же рода вирусов, имеющих сегментированный геном, удается дифференцировать, используя метод молекулярной гибридизации, при к-ром гибридизация РНК-рекомбинанта, обладающего фенотипом вируса дикого типа, с обоими родителями (мутантом и вирусом дикого типа) позволяет четко картировать сегмент РНК, содержащий мутацию. Таким путем были локализованы мутации и определены белки, кодируемые большинством сегментов геномной РНК, у реовирусов, ортомиксовирусов, буньявирусов, аренавирусов, ротавирусов.
В опытах с ДНК-co держащими вирусами позвоночных для картирования мутаций и построения физических и генетических карт с успехом используют фрагменты геномной ДНК, полученные при обработке последней рестрикционными эндонуклеазами, способными разрывать ДНК в определенных участках, характерных для каждой индивидуальной рестриктазы, в результате чего удается получать набор фрагментов (рестриктов) ДНК. При картировании мутаций определяют ре-стрикт ДНК-вируса дикого типа, способный заменить в инфицированных клетках мутантный ген, в результате чего образуется вирус, к-рый в отличие от исходного мутанта может размножаться в неблагоприятных для этого условиях («спасение» мутанта). Поскольку для большинства ДНК-co держащих вирусов построены физические карты геномов, в к-рых определены позиции каждого из рестриктов, получаемых при обработке различными рестриктазами, метод «спасения» мутантов рестриктами ДНК вируса дикого типа позволяет точно локализовать мутацию в геноме и создавать генетические карты. Этот способ можно применять при анализе мутаций паповавирусов, аденовирусов, вирусов герпеса и ортопокс-вирусов. В наст, время разработаны методы анализа отдельных генов вирусов позвоночных — молекулярная гибридизация, пептидное и о лигонуклеотидное картирование, использование моноклональных антител. Они позволяют выявить точечные мутационные изменения в генах вирусов, у к-рых каких-либо фенотипических проявлений мутаций не обнаруживается.
При изучении изменчивости вирусов в природе особое внимание обращают на популяционную изменчивость, т. е. изменчивость не единичных вирионов, а всей вирусной популяции в целом. В основе популяционной изменчивости вирусов лежит первоначальное изменение наследственного материала индивидуальной вирусной частицы, к-рое может быть обусловлено мутацией или рекомбинацией с последующей селекцией тех изменивших наследственные свойства вирионов, к-рые оказались наиболее приспособленными к изменившимся условиям окружающей среды. В результате возникает популяция вируса, обладающая новыми наследственными свойствами. Примером такой изменчивости является возникновение популяций вирусов, устойчивых к противовирусным химиотерапевтическим препаратам, нашедшим достаточно широкое применение, а также появление новых эпидемических и пандемических штаммов вируса гриппа, способных преодолевать иммунитет к ранее циркулировавшим в популяции людей штаммам этого вируса. С помощью новых методов, позволяющих анализировать отдельные гены вирусов, было показано, что у людей могут одновременно циркулировать вирусы, принадлежащие к одному и тому же подтипу, но отличающиеся по ряду генов, причем, как это обнаружено для вируса гриппа, во время одной и той же эпидемии могут циркулировать вирусы, отличающиеся как по антигенной специфичности гемагглютинина, так и по генам, кодирующим внутренние белки вириона. Изучение взаимосвязи между функцией отдельных генов и патогенностью вирусов позвоночных для человека показало, что патогенность является комплексным свойством, в проявлении к-рого участвуют продукты нескольких вирусных генов. Возможность получения патогенных рекомбинантов при скрещивании непатогенных исходных штаммов, так же как и непатогенных рекомбинантов при скрещивании патогенных вирусов, свидетельствует в пользу того, что для проявления патогенности важны не только особенности индивидуальных генов, но и их определенное сочетание в составе генома.
Усовершенствование методов анализа генетических взаимоотношений между вирусами позволило выявить наличие рекомбинации между ts-мутантами пикорнавирусов (вирусы полиомиелита и ящура), хотя частота этой рекомбинации невелика. В результате опытов с РНК-содержащими вирусами, геном к-рых сегментирован, показано, что в основе рекомбинации между этими вирусами, достигающей очень высокой частоты, лежит обмен и перераспределение сегментов генома в процессе совместного культирования скрещиваемых партнеров; в то же время обнаружено, что у этих вирусов может происходить внутригенная рекомбинация по классическому механизму (кроссинговер), частота к-рой, однако, весьма низка. Показана возможность рекомбинации между ортомик-совирусами типа А человека и животных, что имеет особое значение для понимания механизмов возникновения эпидемических штаммов вируса гриппа. У вирусов позвоночных с сегментированным геномом, в частности реовирусов и ортомиксовирусов, в 1979 г. Рамиг и Филдс (R. Ramig, В. Fields) обнаружили феномен, получивший название экст-рагенной супрессии. Суть феномена заключается в том, что продукт определенного сегмента генома, унаследованного рекомбинантом, способен влиять на функцию другого гена, входящего в состав генома рекомбинанта, напр, супрессировать проявление ts-фенотипа. Т. о., при анализе свойств рекомбинантов вирусов позвоночных, особенно обладающих сегментированным геномом, необходимо учитывать, что измененные свойства рекомбинантов могут быть обусловлены не только непосредственно функцией продукта гена, унаследованного от одного из родителей, но также влиянием продукта одного из унаследованных генов на функцию других генов, входящих в состав генома.
Успехи в изучении процессов рекомбинации вирусов позвоночных позволили использовать эти результаты для получения вакцинных вирусных штаммов, в частности вируса гриппа. Так, разработан метод получения высокопродуктивных рекомбинантных вакцинных штаммов для инактивированных вакцин путем рекомбинации высокопродуктивного штамма-донора с новыми антигенными вариантами эпидемических штаммов вируса гриппа. Предложены также способы быстрого получения рекомбинантных вакцинных штаммов для живых гриппозных вакцин. При этом вирус — холодоадаптированный донор рекомбинируют с новыми антигенными вариантами эпидемических штаммов вируса гриппа. Рекомбинанты, унаследовавшие от холодоадаптированного донора 5—6 генов, кодирующих внутренние белки, а от нового эпидемического штамма гемагглютинин и нейраминидазу, после соответствующей проверки используют для приготовления живых вакцин.
В последнее время ведутся исследования по созданию вакцин с помощью методов генетической инженерии. При этом получают клоны кишечной палочки или дрожжей, содержащие соответствующую ДНК-копию вирусного гена, кодирующего белок, ответственный за иммунитет. Этот белок, продуцируемый бактериальным клоном, предполагают использовать в качестве инактивированной вакцины. Другой метод заключается в введении ответственного за иммунитет гена одного вируса в геном крупного ДНК-со-держащего вируса (напр., вируса осповакцины), имеющего делецию определенной области генома, не являющейся жизненно важной для вируса (вирус-вектор). Такой гибридный вирус, размножаясь в вакцинированном организме, может вызвать иммунитет и к тому чужеродному вирусу, ген к-рого содержится в геноме этого вируса-вектора. Следует, однако, отметить, что пока вирусные вакцины, получаемые с помощью методов генетической инженерии, не нашли практического применения, хотя исследования в этом направлении ведутся во многих странах.
Вирусы беспозвоночных
составляют большую группу В., поражающих гл. обр. представителей класса насекомых (Insecta). В последнее время стали известны также вирусные заболевания наукообразных клещей (класс Arachnoidea) и ракообразных (класс Crustacea). Имеются сообщения о наличии вирусоподобных частиц у амебы и парамеций (Protozoa).
В организме насекомых В. проходят полный цикл своего развития. Согласно единой классификации В. насекомых образуют семь групп, две из которых имеют статус рода (Baculovirus и Iridovirus). Наиболее обширным родом является род Baculovirus; вирионы имеют бациллярную форму и размеры 250—400 X Х 40—70 нм, внутреннюю и внешнюю мембраны и содержат двунитчатую ДНК с мол. весом ок. 80Х10^6. К подгруппе «А» этого рода относятся В. полиэдрозов, образующие в клетках тканей больного насекомого кристаллические белковые включения, полиэдры, содержащие несколько десятков вирионов. К подгруппе «В» относятся вирусы гранулезов, образующие гранулы или капсулы, содержащие один, реже два вириона. Средние размеры полиэдров 2—4 мкм, средние размеры гранул 200—400 нм. Представители рода Baculovirus поражают наибольшее число насекомых из отряда чешуекрылых (Lepidoptera), меньше перепончатокрылых (Hymenoptera) и другие отряды насекомых. Типичным представителем этого рода является В. полиэдроза тутового шелкопряда (Bombyx mori), приводящий к гибели гусениц на промышленных выкормах. Характерной особенностью этого В. является образование в ядрах пораженных клеток многочисленных шестигранных полиэдров размером 3—4 мкм, устойчивых к растворению в воде, растворителям липидов, бактериальному разложению. Полиэдры растворяются в щелочах и кислотах. Симптомы заболевания: вялость гусениц, пожелтение покровов, раздутость тела, мутно-молочная гемолимфа. Поражаются гиподерма, жировое тело, трахеальный матрикс, гемолимфа. Гибель гусениц наступает на 7—8-й день после инфицирования с лизисом всех тканей. Представителем В. гранулезов может служить В. гранулеза хвоевертки, Choristoneura murinana. Его характерной особенностью является образование многочисленных гранул в цитоплазме пораженных клеток. Симптомы заболевания и поражаемые ткани сходны с полиэдрозами. Род Iridovirus представлен «радужными» В., характеризующимися образованием радужного свечения зелено-голубых, фиолетовых или оранжевых тонов в пораженном насекомом. Эффект радужности зависит от дифракции видимого света чрезвычайно правильной, паракристаллической упаковкой вирусных частиц. В. локализуется в цитоплазме клеток жирового тела. Вирионы имеют сферическую форму с диам. 130 нм и внешнюю оболочку, состоящую из 1500 капсомеров. В состав вирионов входит 15% двунитчатой ДНК с мол. весом ок. 140Х10 6 .
Группа В. цитоплазменного полиэдроза относится к сем. Reoviridae. Вирионы с диам. 60 нм представлены икосаэдрами и содержат 21—23% двунитчатой РНК. Они заключены в крупные полиэдры и локализуются в цитоплазме клеток средней кишки пораженного насекомого. Поражаются насекомые из отряда чешуекрылых и сетчатокрылых (Neuroptera). Трупы погибших гусениц затвердевают, кишечник приобретает беловатый оттенок. Типичный представитель — В. цитоплазменного полиэдроза тутового шелкопряда. РНК этого В. имеет мол. вес 12,7—21 X 10 6 и состоит из двух фрагментов — 12S и 15S.
К сем. Poxviridae относятся «энтомопоксвирусы». Они содержат ДНК, локализуются в цитоплазме и имеют овальные вирионы размером 300— 400 нм. Вирионы заключены в веретенообразные или яйцевидные включения (до 15 мкм). Характерна ребристая поверхность вирионов. Типичный представитель — В. майского хруща (Melolontha melolontha).
К роду Parvovirus отнесен В. денсонуклеоза, или болезнь плотных ядер — Densonucleosis virus Galleria. В. образует изометрические частицы, икосаэдральной симметрии с диам. 21—23 нм и содержит 30% однонитчатой ДНК с мол. весом 4—5 Х 10 6 . Описан у большой пчелиной моли (Galleria melonella), у к-рой он вызывает паралич и смерть. Характерно образование плотных фейльгенположительных масс в сильно гипертрофированных ядрах жирового тела. В. высокоспецифичен.
Одна из групп В. насекомых близка по свойствам к энтеровирусам. Представителем этой группы является В. острого паралича пчел. Вирионы — изометрические сферические частицы с диам. 20—30 нм, содержат однонитчатую РНК и локализуются в цитоплазме клеток жирового тела. Паралич и гибель пчел наступают через 2—4 дня после инфицирования.
В. сигма дрозофилы представляет группу В., близких к В. везикулярного стоматита. Вирионы бациллярной формы с закруглением на одном конце с диам. 70 X 140 нм, предположительно РНК-типа. Характеризуется чувствительностью пораженных В. мух, их личинок и куколок к углекислоте. При 20—40% концентрации углекислоты в атмосфере наступает паралич и смерть мух. В. передается трансовариально, а также через сперму.
В. насекомых широко распространены в природе. Из 300 В. насекомых ок. 200 принадлежат к В. полиэдрозов и гранулезов, остальные — к В., не образующим включений. Характерно латентное вирусоносительство. Активация латентного В. под действием различных стрессоров приводит к эпизоотиям среди вредных и полезных насекомых, снижая их численность. Поражаются гл. обр. личиночные стадии насекомых. Заражение происходит через пищу, поврежденные покровы, трансовариально. Инфекционным материалом служат трупы погибших гусениц, загрязненная ими пища, фекалии, гемолимфа. В. могут распространяться больными гусеницами, ветром, водой, через почву, птицами. Инкубационный период зависит от дозы В., возраста гусениц и внешних факторов (особенно температуры). В пораженном насекомом В. накапливается в количестве 10—30% от веса тканей. Чаще наблюдается групповая специфичность: один В. поражает обычно близкие виды, но некоторые В. могут поражать различные виды, роды и даже отряды насекомых. Так, радужный В. Tipula экспериментально передан 7 видам двукрылых (Diptera), И видам чешуекрылых (Lepidoptera) и 3 видам жесткокрылых (Coleoptera).
Благодаря значительному накоплению В. в теле гусениц, образованию эпизоотий и безвредности для человека и животных В. беспозвоночных используются в биол, борьбе с вредными насекомыми. Они используются также как чрезвычайно удобные модели для исследования вопросов общей вирусологии.
Вирусы растений
По морфологии вирионов В. растений относятся к четырем группам: 1. Спиральные палочковидные В., имеющие жесткую структуру, напр. ВТМ (300X18 нм), В. штриховатой мозаики ячменя (130X19 нм). 2. Спиральные нитевидные В., отличающиеся значительной длиной и эластичностью вирионов, напр. Х-вирус картофеля (520 X 12 нм), В. желтухи сахарной свеклы (1250X 10 нм), В. мозаики плевела (1725X18 нм). 3. Сферические или икосаэдрические В., напр. В. некроза табака (20 нм), желтой мозаики турнепса (30 нм). 4. Бацилловидные или пулевидные В., содержащие мембраны и внутренний нуклеокапсид, напр. В. карликовости кукурузы (240X48 нм), желтой карликовости картофеля (380X75 нм), мозаики озимой пшеницы (260X60 нм). Структура жестких палочек известна только у В. растений, нитевидные формы, несущие РНК, также найдены только у В. растений, сферические вирионы известны у бактериофагов и В. животных, бацилловидные — у В. животных.
Представитель первой группы — ВТМ — имеет вирион в форме жесткого цилиндра с размерами 300 X 18 нм и мол. весом 39 X 106 дальтон. Стенка цилиндра образована спирально уложенными одинаковыми белковыми молекулами — субъединицами, число которых составляет ок. 2130. Мол. вес субъединицы ок. 18 000 дальтон. Витки спирали плотно прилегают друг к другу и каждый из них включает 161/3 субъединиц.
Тяж вирусной РНК, состоящий приблизительно из 6400 нуклеотидов и имеющий мол. вес 2Х10 6 дальтон, проходит между рядами белковых субъединиц, следуя их спиральному расположению, на расстоянии 4 нм от центра поперечного сечения частицы. При таком расположении на каждые три нуклеотида тяжа РНК приходится одна белковая субъединица. Внутри цилиндра, по всей его длине, проходит полость с диам. 4 нм. Вирусная РНК защищена белком от действия клеточных нуклеаз, но низкомолекулярные вещества, в т. ч. мутагены, могут проникать в область локализации РНК, оказывая на нее инактивирующее или мутагенное действие. В экспериментальных условиях вирионы некоторых палочковидных и сферических В. растений удалось дезагрегировать на белковые субъединицы и РНК с сохранением их нативного состояния и затем вновь вызвать агрегацию этих компонентов с восстановлением структуры, инфекционности и других свойств вирионов.
У большинства В. растений геном представлен однотяжной РНК, но В. раневых опухолей растений и карликовости риса имеют двутяжную РНК. ДНК найдена пока только у сферического вируса мозаики цветной капусты. Известны В. растений, РНК-геном которых фрагментирован, каждый из фрагментов несет часть генетической информации и заключен в отдельный капсид. Только объединенное действие всех фрагментов обеспечивает функции полного генома. К таким В. растений относятся возбудители погремковости табака, мозаики люцерны и др. У некоторых В. растений геном настолько мал, что не обеспечивает репликацию вирусной РНК. Так, у сателлита В. некроза табака мол. вес РНК составляет всего лишь 0,4х10^6 дальтон, он индуцирует синтез собственного оболочечного белка, но не имеет цистрона синтетазы и паразитирует за счет неродственного В. некроза табака, используя его синтетазу.
В. растений проникают в клетки растений путем пиноцитоза (см.) при повреждениях, обнажающих участки плазматической мембраны. В природе они распространяются гл. обр. сосущими насекомыми, клещами, нематодами и зооспорами некоторых низших грибов — корневых паразитов. Известны В. растений, размножающиеся в организме насекомых-переносчиков, часть из них передается потомству переносчиков через яйцо. Некоторые В. распространяются семенами больных растений. Вегетативное размножение растений особенно способствует распространению В.
Внутриклеточное развитие В. растений следует тем же закономерностям, какие известны для репродукции РНК-содержащих В. вообще. Многие В. растений образуют в клетках кристаллические включения, состоящие из вирионов, и аморфные, содержащие В. и компоненты цитоплазмы. Болезни растений, вызванные В., хронические, выздоровления не наступает. Интенсивное размножение В. происходит в растущих листьях и других органах. В меристеме точки роста В. немного, в апикальной ее части он может отсутствовать, на этом основан прием стерильного выращивания здоровых растений из кусочков меристемы. Некоторые В. инактивируются в зараженных растениях при тепловой обработке. Химиотерапия против В. растений не разработана.
Одни В. поражают растения многих видов, другие узко специализированы. ВТМ, напр., в опытных условиях способен заразить растения многих видов типа цветковых, нескольких видов типа папоротниковидных, он инфицирует протопласты дрожжей, а его РНК реплицируется и транслируется in vitro даже во фракции митохондрий, изолированной из клеток печени крысы.
Отдельные группы вирусов — см. статьи по названию группы вирусов (напр., Аденовирусы, Ареновирусы, Кишечные вирусы, Онкогенные вирусы, Парвовирусы и др.).
https://bio-lessons.ru/virusy-bakteriofagi-virusnye-zabolevanija/
https://xn--90aw5c.xn--c1avg/index.php/%D0%92%D0%98%D0%A0%D0%A3%D0%A1%D0%AB